

Temperature-mediated flower size plasticity in Arabidopsis
- PMID: 36388994
- PMCID: PMC9646949
- DOI: 10.1016/j.isci.2022.105411
Temperature-mediated flower size plasticity in Arabidopsis
Abstract
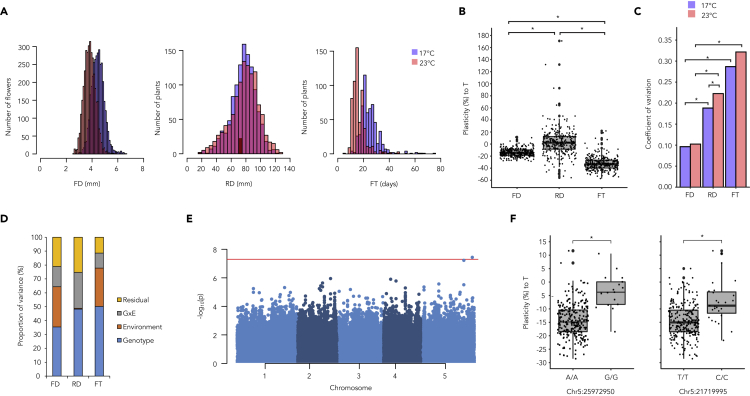
Organisms can rapidly mitigate the effects of environmental changes by changing their phenotypes, known as phenotypic plasticity. Yet, little is known about the temperature-mediated plasticity of traits that are directly linked to plant fitness such as flower size. We discovered substantial genetic variation in flower size plasticity to temperature both among selfing Arabidopsis thaliana and outcrossing A. arenosa individuals collected from a natural growth habitat. Genetic analysis using a panel of 290 A. thaliana accession and mutant lines revealed that MADS AFFECTING FLOWERING (MAF) 2-5 gene cluster, previously shown to regulate temperature-mediated flowering time, was associated to the flower size plasticity to temperature. Furthermore, our findings pointed that the control of plasticity differs from control of the trait itself. Altogether, our study advances the understanding of genetic and molecular factors underlying plasticity on fundamental fitness traits, such as flower size, in response to future climate scenarios.
Keywords: Plant biology; Plant genetics; Plant morphology; Plant physiology.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Alseekh S., Aharoni A., Brotman Y., Contrepois K., D'Auria J., Ewald J., Ewald J.C., Fraser P.D., Giavalisco P., Hall R.D., et al. Mass spectrometry-based metabolomics: a guide for annotation, quantification and best reporting practices. Nat. Methods. 2021;18:747–756. doi: 10.1038/s41592-021-01197-1. - DOI - PMC - PubMed
-
- Amiri S., Zwanzig S. Assessing the coefficient of variations of chemical data using bootstrap method. J. Chemom. 2011;25:295–300. doi: 10.1002/cem.1350. - DOI
-
- Barnes A.C., Rodríguez-Zapata F., Juárez-Núñez K.A., Gates D.J., Janzen G.M., Kur A., Wang L., Jensen S.E., Estévez-Palmas J.M., Crow T.M., et al. An adaptive teosinte mexicana introgression modulates phosphatidylcholine levels and is associated with maize flowering time. Proc. Natl. Acad. Sci. USA. 2022;119 doi: 10.1073/pnas.2100036119. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
