

Finding memo: versatile interactions of the VPS10p-Domain receptors in Alzheimer's disease
- PMID: 36397124
- PMCID: PMC9673319
- DOI: 10.1186/s13024-022-00576-2
Finding memo: versatile interactions of the VPS10p-Domain receptors in Alzheimer's disease
Abstract
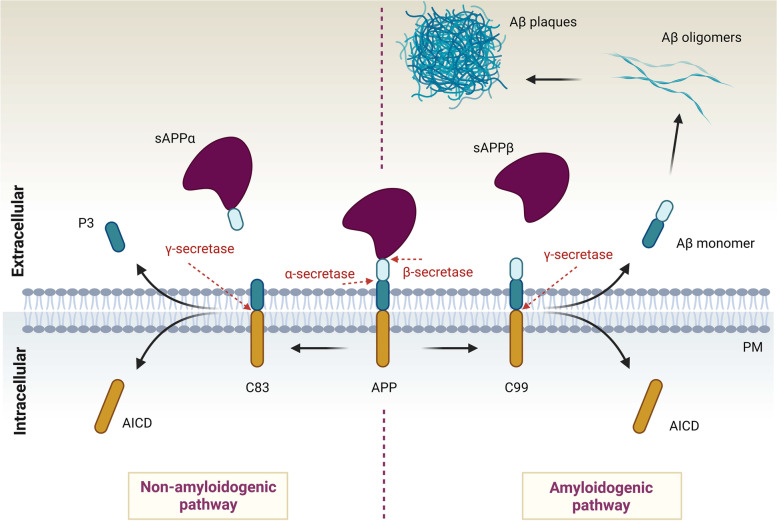
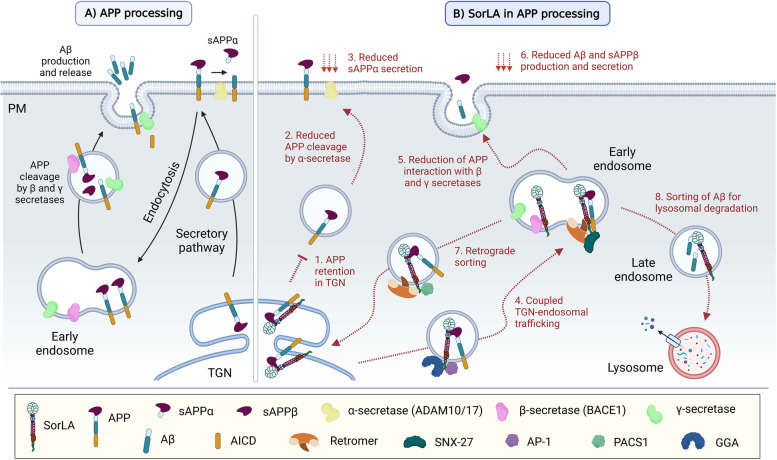
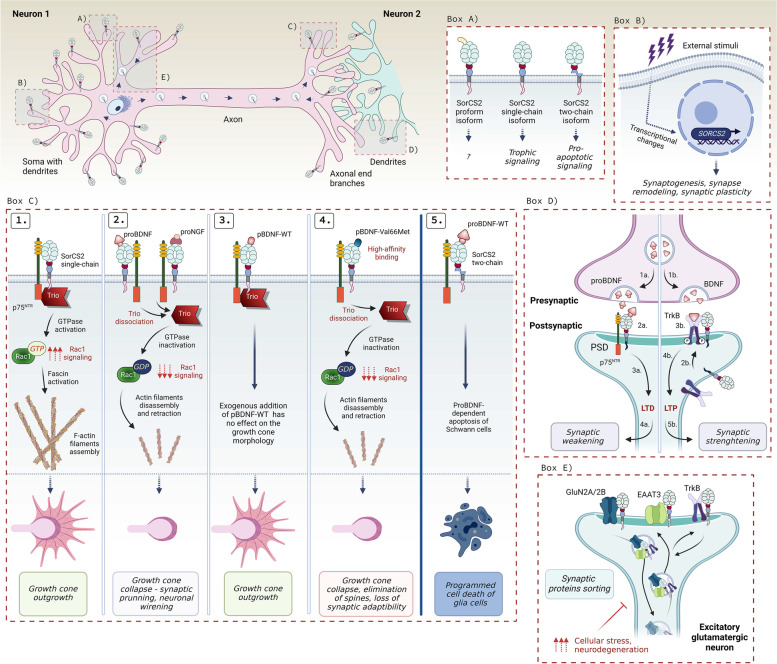
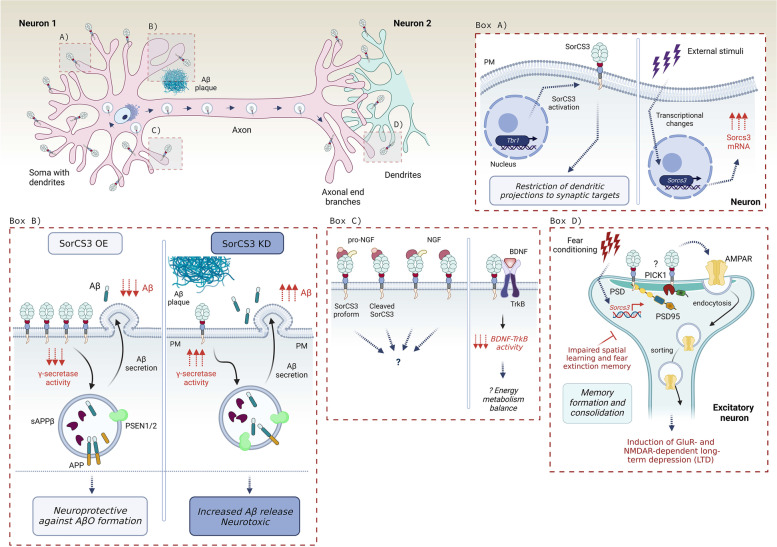
The family of VPS10p-Domain (D) receptors comprises five members named SorLA, Sortilin, SorCS1, SorCS2 and SorCS3. While their physiological roles remain incompletely resolved, they have been recognized for their signaling engagements and trafficking abilities, navigating a number of molecules between endosome, Golgi compartments, and the cell surface. Strikingly, recent studies connected all the VPS10p-D receptors to Alzheimer's disease (AD) development. In addition, they have been also associated with diseases comorbid with AD such as diabetes mellitus and major depressive disorder. This systematic review elaborates on genetic, functional, and mechanistic insights into how dysfunction in VPS10p-D receptors may contribute to AD etiology, AD onset diversity, and AD comorbidities. Starting with their functions in controlling cellular trafficking of amyloid precursor protein and the metabolism of the amyloid beta peptide, we present and exemplify how these receptors, despite being structurally similar, regulate various and distinct cellular events involved in AD. This includes a plethora of signaling crosstalks that impact on neuronal survival, neuronal wiring, neuronal polarity, and synaptic plasticity. Signaling activities of the VPS10p-D receptors are especially linked, but not limited to, the regulation of neuronal fitness and apoptosis via their physical interaction with pro- and mature neurotrophins and their receptors. By compiling the functional versatility of VPS10p-D receptors and their interactions with AD-related pathways, we aim to further propel the AD research towards VPS10p-D receptor family, knowledge that may lead to new diagnostic markers and therapeutic strategies for AD patients.
Keywords: Alzheimer’s disease; Comorbidity; Neurotrophins; SorCS1; SorCS2; SorCS3; SorLA; Sortilin.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
-
- 2020 Alzheimer's disease facts and figures. Alzheimer's & Dementia 2020;16(3):391-460. 10.1002/alz.12068. - PubMed
-
- Wang J, Gu BJ, Masters CL, Wang Y-J. A systemic view of Alzheimer disease — insights from amyloid-β metabolism beyond the brain. Nat Rev Neurol. 2017;13:612. - PubMed
-
- Panza F, Lozupone M, Logroscino G, Imbimbo BP. A critical appraisal of amyloid-β-targeting therapies for Alzheimer disease. Nat Rev Neurol. 2019;15:73–88. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
