

Human pericytes degrade diverse α-synuclein aggregates
- PMID: 36399706
- PMCID: PMC9674377
- DOI: 10.1371/journal.pone.0277658
Human pericytes degrade diverse α-synuclein aggregates
Abstract
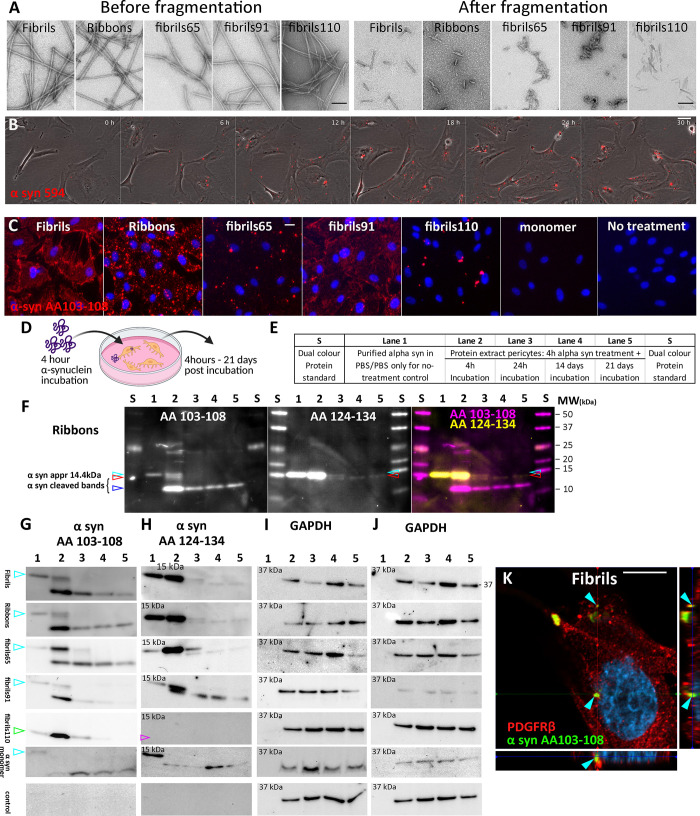
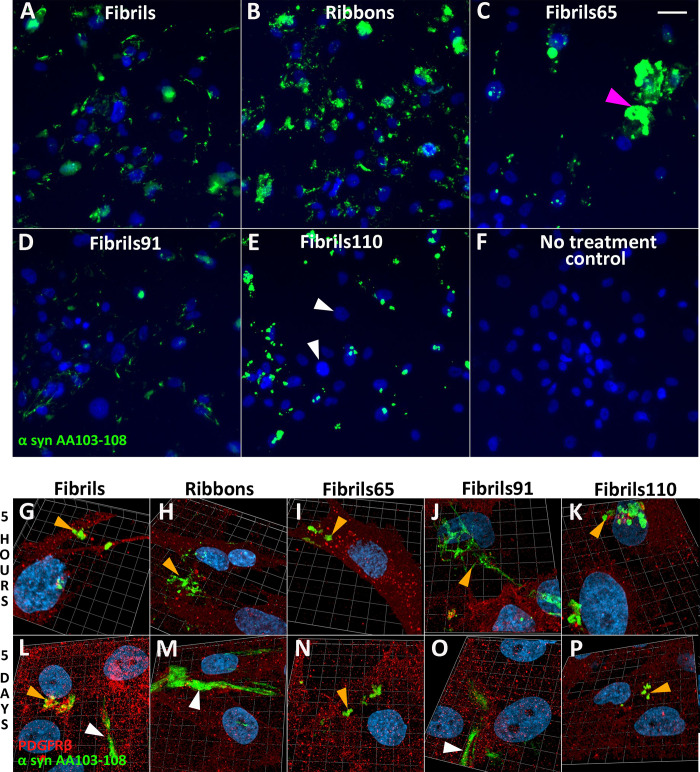
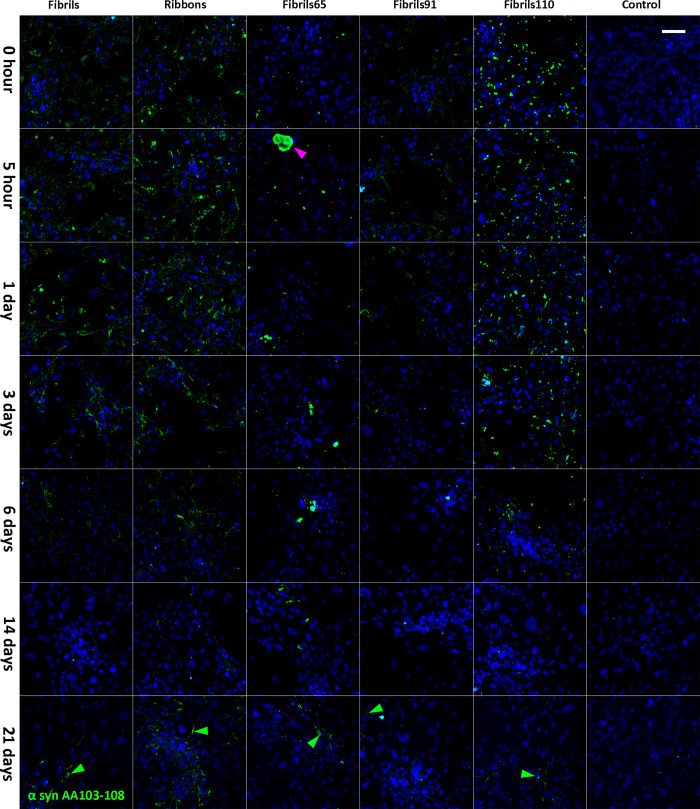
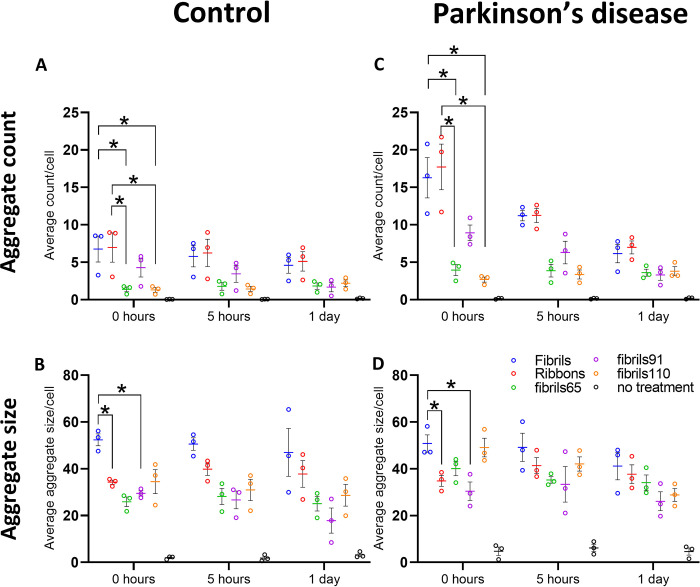
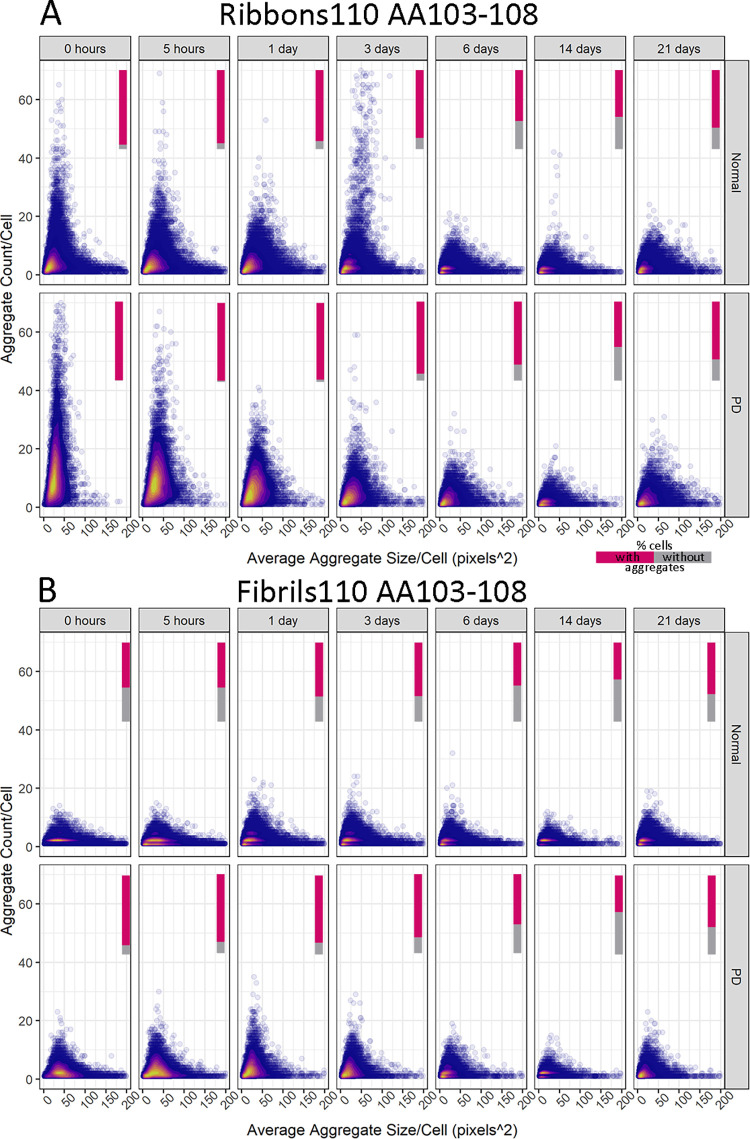
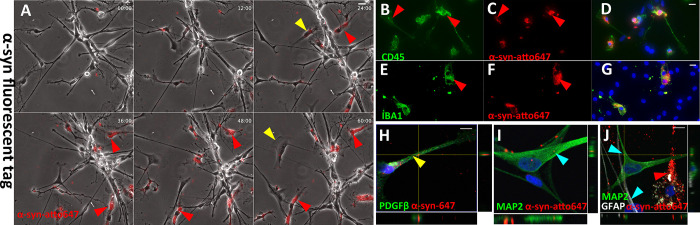
Parkinson's disease (PD) is a progressive, neurodegenerative disorder characterised by the abnormal accumulation of α-synuclein (α-syn) aggregates. Central to disease progression is the gradual spread of pathological α-syn. α-syn aggregation is closely linked to progressive neuron loss. As such, clearance of α-syn aggregates may slow the progression of PD and lead to less severe symptoms. Evidence is increasing that non-neuronal cells play a role in PD and other synucleinopathies such as Lewy body dementia and multiple system atrophy. Our previous work has shown that pericytes-vascular mural cells that regulate the blood-brain barrier-contain α-syn aggregates in human PD brains. Here, we demonstrate that pericytes efficiently internalise fibrillar α-syn irrespective of being in a monoculture or mixed neuronal cell culture. Pericytes cleave fibrillar α-syn aggregates (Fibrils, Ribbons, fibrils65, fibrils91 and fibrils110), with cleaved α-syn remaining present for up to 21 days. The number of α-syn aggregates/cell and average aggregate size depends on the type of strain, but differences disappear within 5 five hours of treatment. Our results highlight the role brain vasculature may play in reducing α-syn aggregate burden in PD.
Copyright: © 2022 Dieriks et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
