

Glucagon receptor antagonism impairs and glucagon receptor agonism enhances triglycerides metabolism in mice
- PMID: 36400402
- PMCID: PMC9706156
- DOI: 10.1016/j.molmet.2022.101639
Glucagon receptor antagonism impairs and glucagon receptor agonism enhances triglycerides metabolism in mice
Abstract
Objective: Treatment with glucagon receptor antagonists (GRAs) reduces blood glucose but causes dyslipidemia and accumulation of fat in the liver. We investigated the acute and chronic effects of glucagon on lipid metabolism in mice.
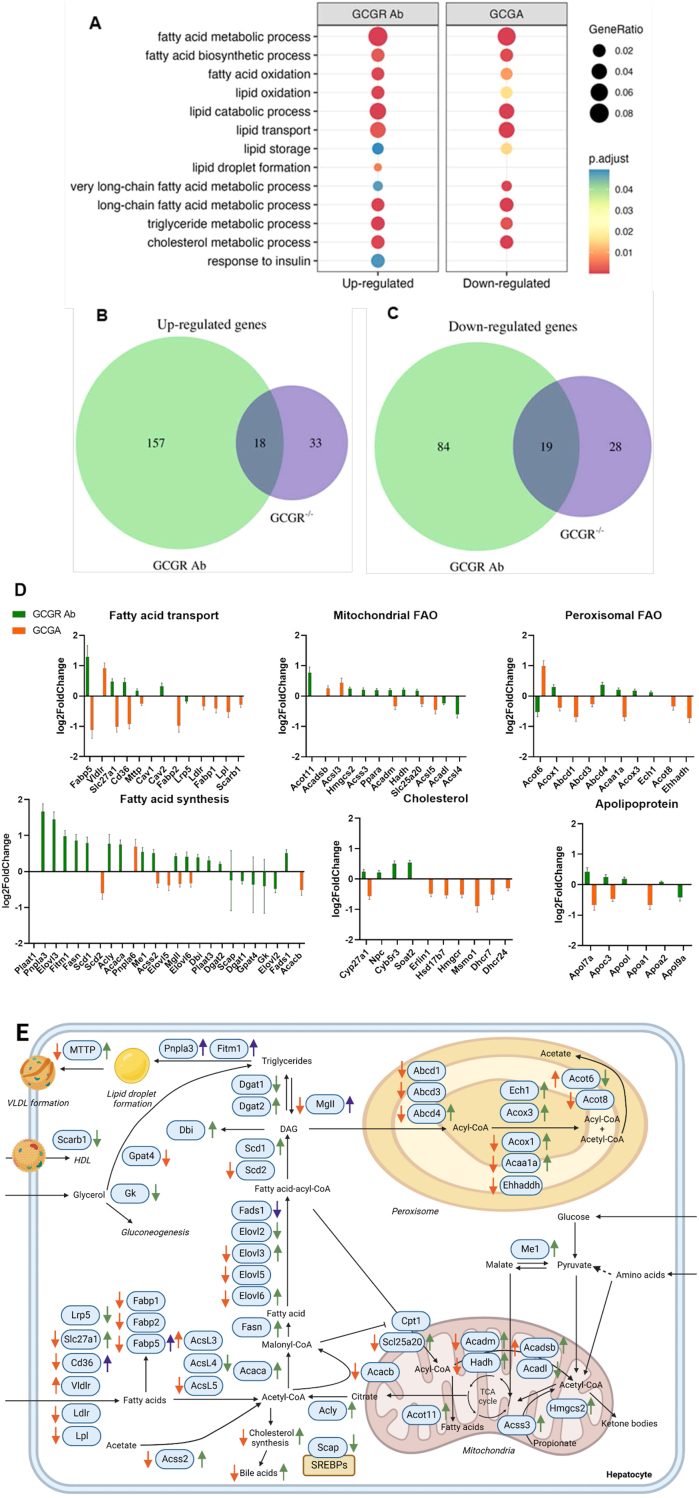
Methods: Chronic effects of glucagon receptor signaling on lipid metabolism were studied using oral lipid tolerance tests (OLTTs) in overnight fasted glucagon receptor knockout (Gcgr-/-) mice, and in C57Bl/6JRj mice treated with a glucagon receptor antibody (GCGR Ab) or a long-acting glucagon analogue (GCGA) for eight weeks. Following treatment, liver tissue was harvested for RNA-sequencing and triglyceride measurements. Acute effects were studied in C57Bl/6JRj mice treated with a GRA or GCGA 1 h or immediately before OLTTs, respectively. Direct effects of glucagon on hepatic lipolysis were studied using isolated perfused mouse liver preparations. To investigate potential effects of GCGA and GRA on gastric emptying, paracetamol was, in separate experiments, administered immediately before OLTTs.
Results: Plasma triglyceride concentrations increased 2-fold in Gcgr-/- mice compared to their wild-type littermates during the OLTT (P = 0.001). Chronic treatment with GCGR Ab increased, whereas GCGA treatment decreased, plasma triglyceride concentrations during OLTTs (P < 0.05). Genes involved in lipid metabolism were upregulated upon GCGR Ab treatment while GCGA treatment had opposite effects. Acute GRA and GCGA treatment, respectively, increased (P = 0.02) and decreased (P = 0.003) plasma triglyceride concentrations during OLTTs. Glucagon stimulated hepatic lipolysis, evident by an increase in free fatty acid concentrations in the effluent from perfused mouse livers. In line with this, GCGR Ab treatment increased, while GCGA treatment decreased, liver triglyceride concentrations. The effects of glucagon appeared independent of changes in gastric emptying of paracetamol.
Conclusions: Glucagon receptor signaling regulates triglyceride metabolism, both chronically and acutely, in mice. These data expand glucagon´s biological role and implicate that intact glucagon signaling is important for lipid metabolism. Glucagon agonism may have beneficial effects on hepatic and peripheral triglyceride metabolism.
Keywords: Cholesterol; Glucagon; Non-esterified/ free fatty acids; Steatosis; Triglycerides.
Copyright © 2022 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures






References
-
- Kazda C.M., Ding Y., Kelly R.P., Garhyan P., Shi C., Lim C.N., et al. Evaluation of efficacy and safety of the glucagon receptor antagonist LY2409021 in patients with type 2 diabetes: 12- and 24-week phase 2 studies. Diabetes Care. 2016;39(7):1241–1249. - PubMed
-
- Kazda C.M., Frias J., Foga I., Cui X., Guzman C.B., Garhyan P., et al. Treatment with the glucagon receptor antagonist LY2409021 increases ambulatory blood pressure in patients with type 2 diabetes. Diabetes, Obesity and Metabolism. 2017;19(8):1071–1077. - PubMed
-
- Guzman C.B., Zhang X.M., Liu R., Regev A., Shankar S., Garhyan P., et al. Treatment with LY2409021, a glucagon receptor antagonist, increases liver fat in patients with type 2 diabetes. Diabetes, Obesity and Metabolism. 2017;19(11):1521–1528. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
