

Preferential Modulatory Action of 5-HT2A Receptors on the Dynamic Regulation of Basal Ganglia Circuits
- PMID: 36400530
- PMCID: PMC9838704
- DOI: 10.1523/JNEUROSCI.1181-22.2022
Preferential Modulatory Action of 5-HT2A Receptors on the Dynamic Regulation of Basal Ganglia Circuits
Erratum in
-
Erratum: Guilhemsang et al., "Preferential Modulatory Action of 5-HT2A Receptors on the Dynamic Regulation of Basal Ganglia Circuits".J Neurosci. 2023 Mar 15;43(11):2033-2034. doi: 10.1523/JNEUROSCI.0224-23.2023. Epub 2023 Feb 23. J Neurosci. 2023. PMID: 36823041 Free PMC article. No abstract available.
Abstract
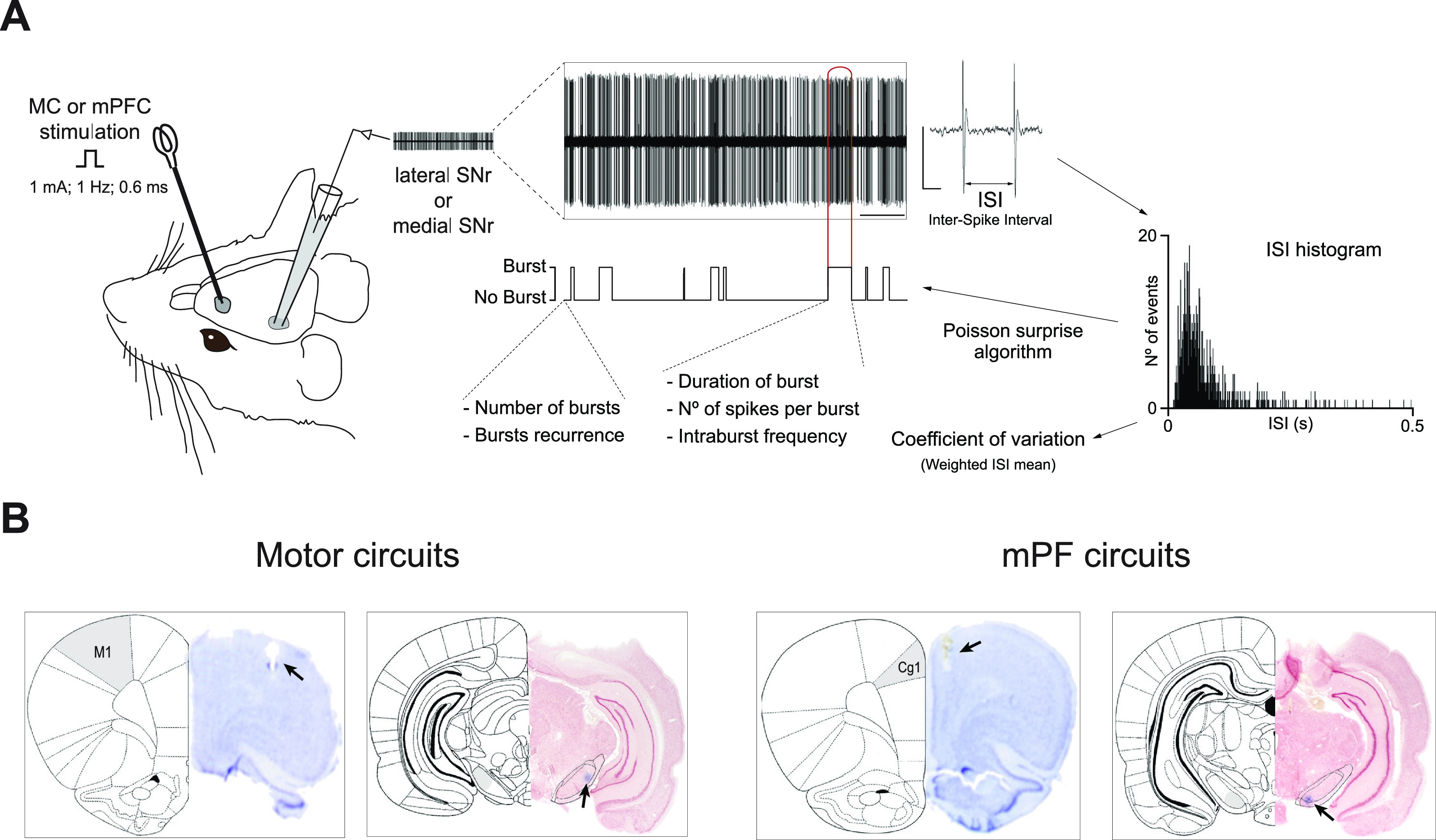
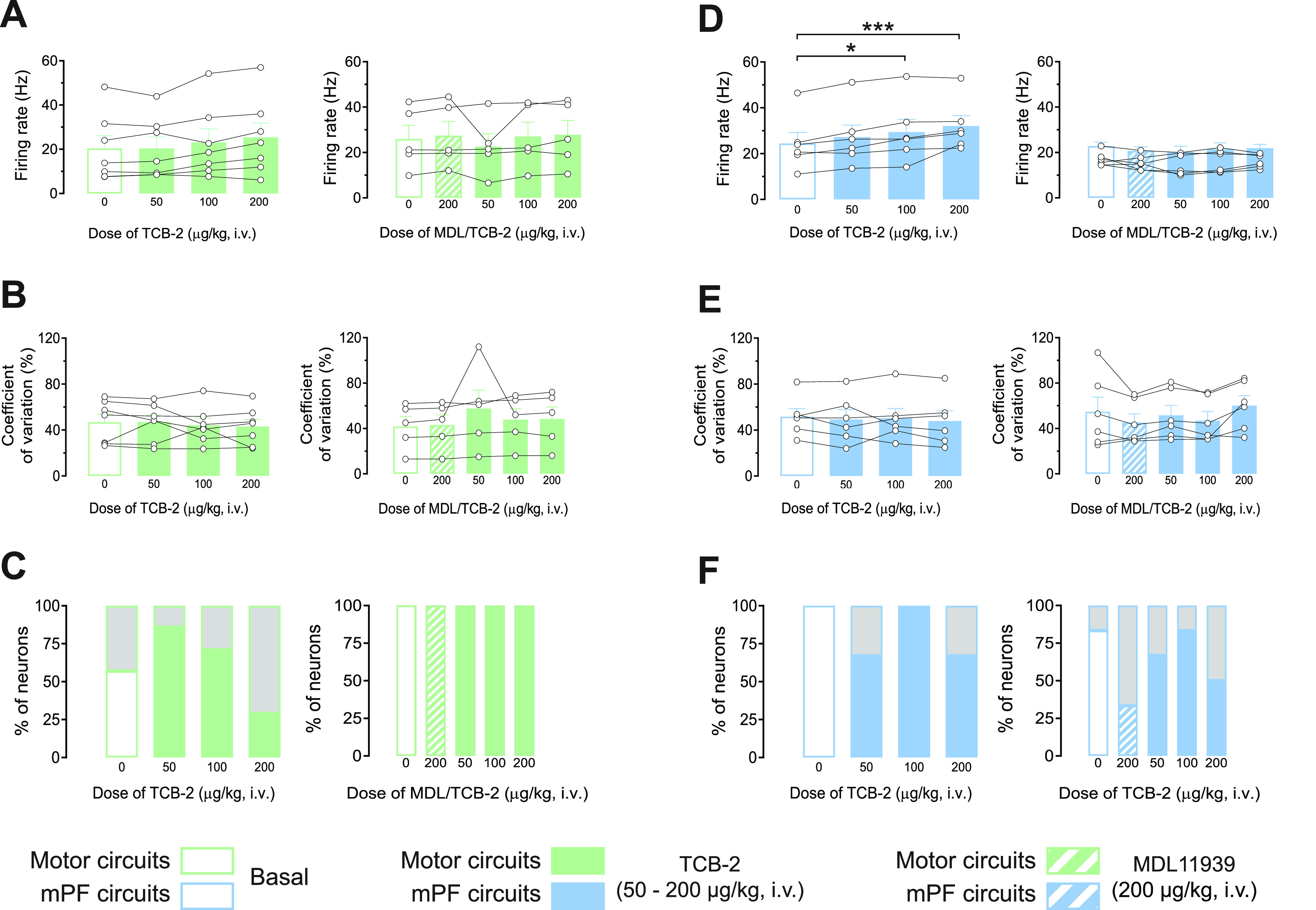
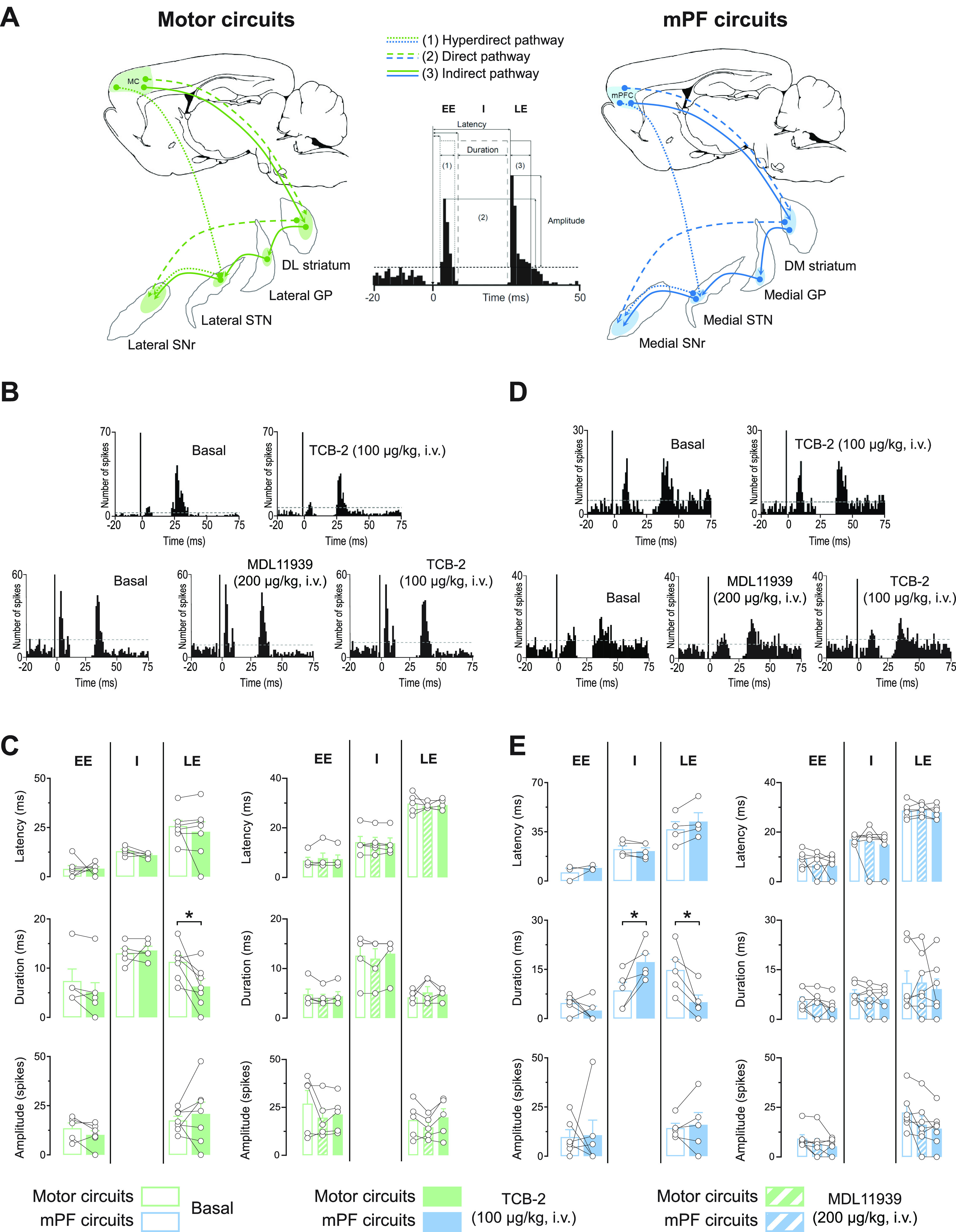
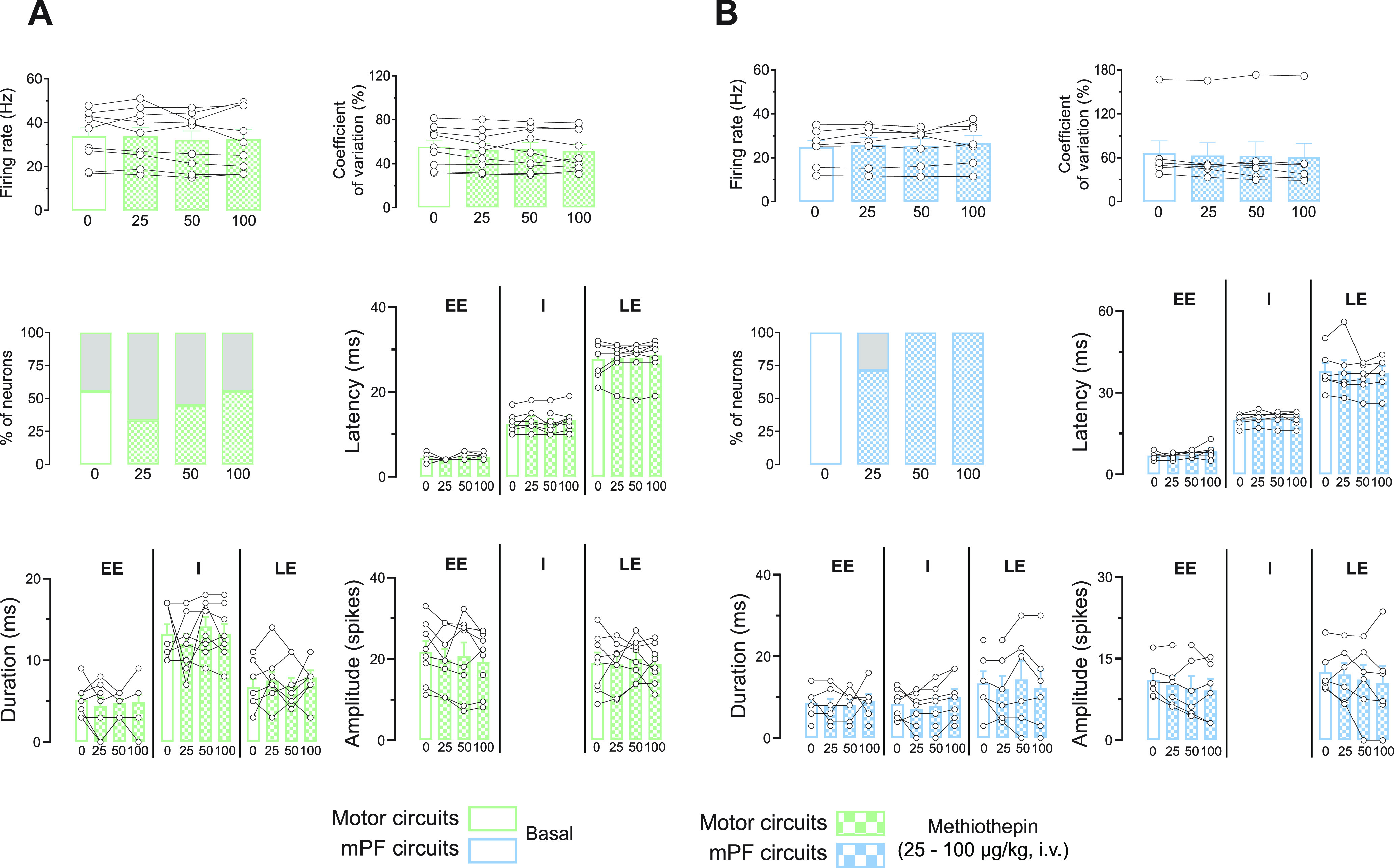
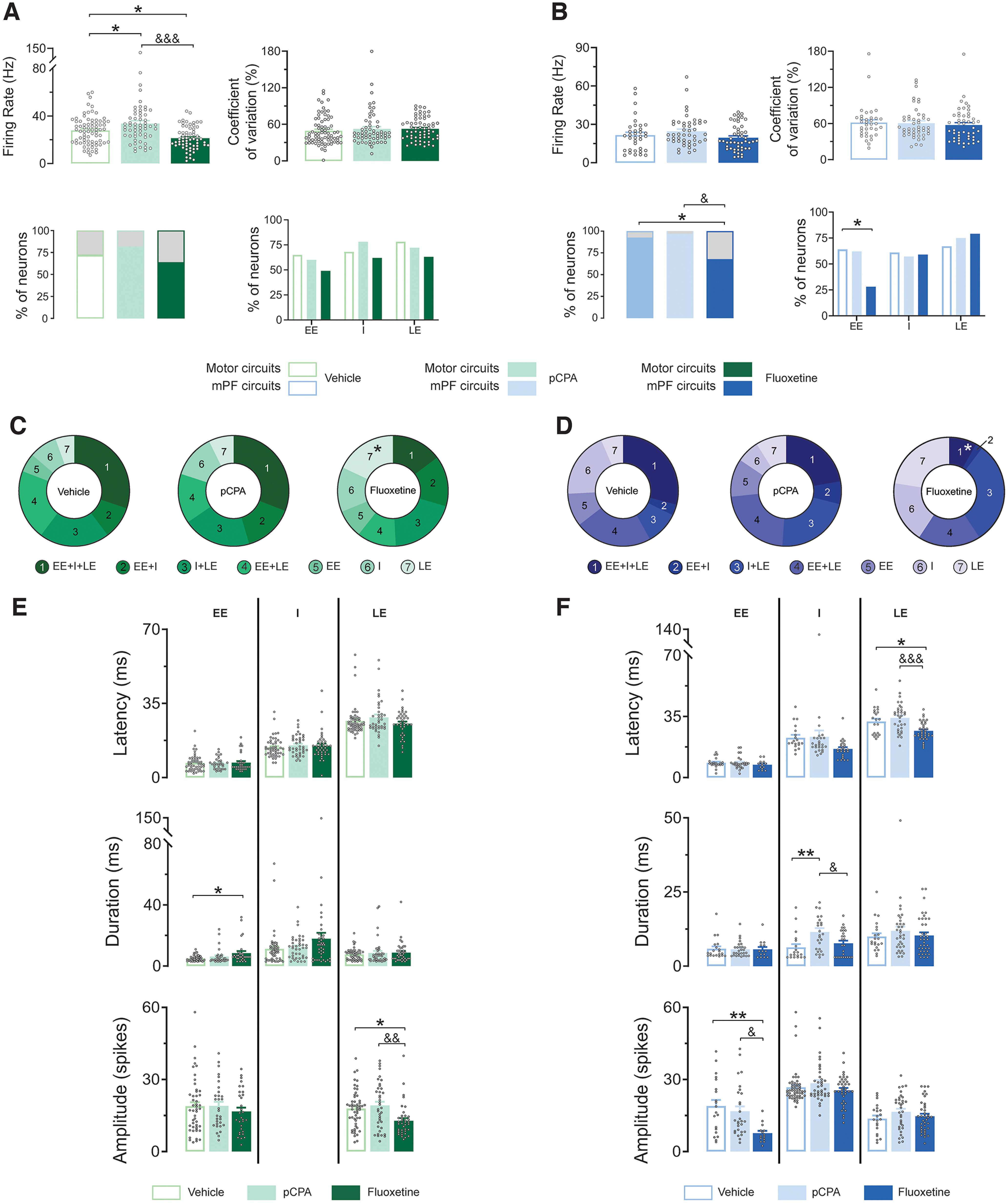
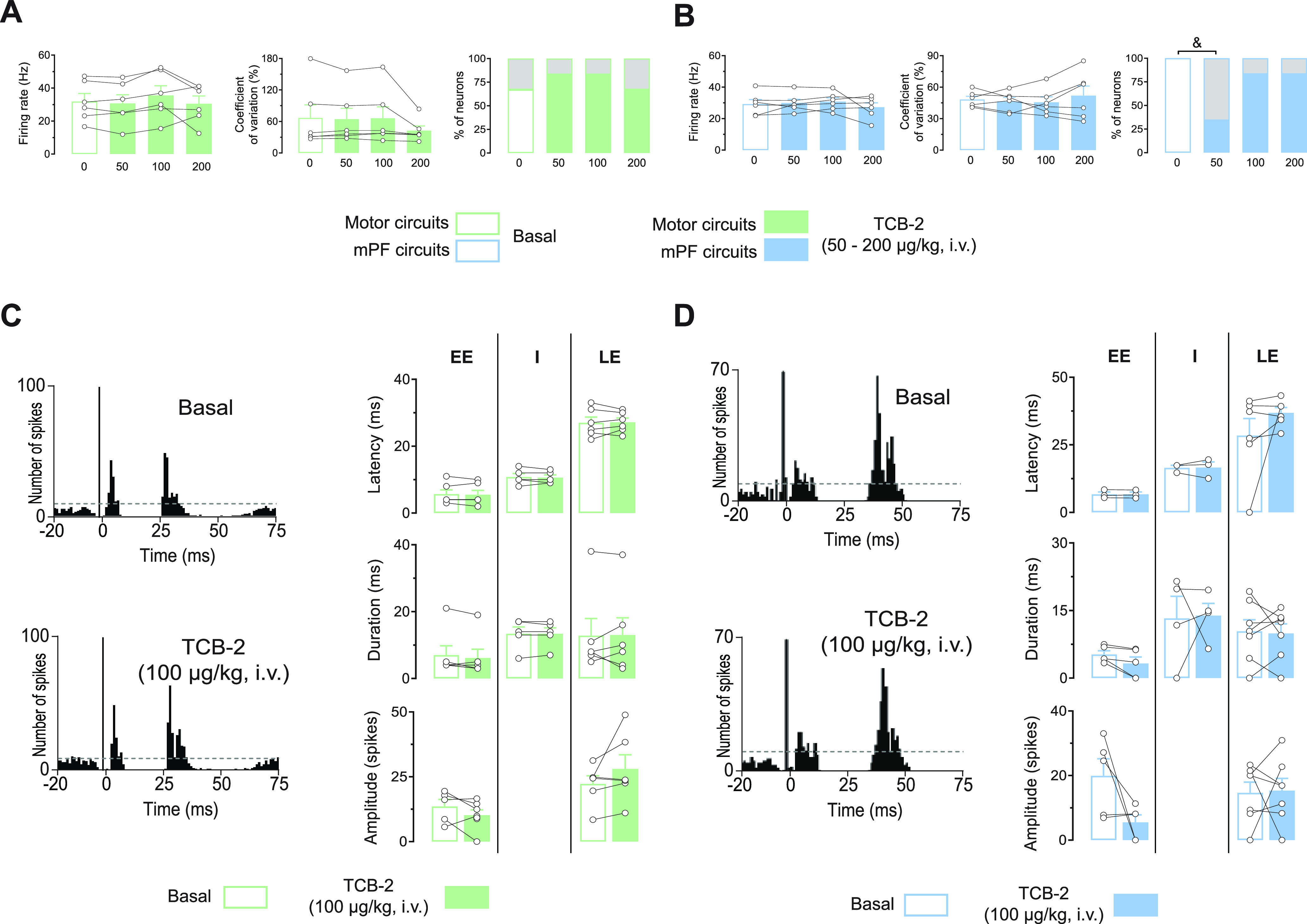
In rodents, cortical information is transferred to the substantia nigra pars reticulata (SNr) through motor and medial prefrontal (mPF) basal ganglia (BG) circuits implicated in motor and cognitive/motivational behaviors, respectively. The serotonergic 5-HT2A receptors are located in both of these neuronal networks, displaying topographical differences with a high expression in the associative/limbic territories, and a very low expression in the subthalamic nucleus. This study investigated whether the stimulation of 5-HT2A receptors could have a specific signature on the dynamic regulation of BG circuits, preferentially modulating the mPF information processing through trans-striatal pathways. We performed in vivo single-unit extracellular recordings to assess the effect of the 5-HT2A agonist TCB-2 on the spontaneous and cortically evoked activity of lateral and medial SNr neurons in male rats (involved in motor and mPF circuits, respectively). TCB-2 (50-200 µg/kg, i.v.) increased the basal firing rate and enhanced the cortically evoked inhibitory response of medial SNr neurons (transmission through the direct striato-nigral pathway). A prior administration of the preferential 5-HT2A receptor antagonist MDL11939 (200 µg/kg, i.v.) did not modify any electrophysiological parameter, but occluded TCB-2-induced effects. In animals treated with the 5-HT synthesis inhibitor pCPA (4-chloro-dl-phenylalanine methyl ester hydrochloride), TCB-2 failed to induce the above-mentioned effects, thus suggesting the contribution of endogenous 5-HT. However, the mobilization of 5-HT induced by the acute administration of fluoxetine (10 mg/kg, i.p.) did not mimic the effects triggered by TCB-2. Overall, these data suggest that 5-HT2A receptors have a preferential modulatory action on the dynamic regulation of BG circuitry.SIGNIFICANCE STATEMENT Motor and medial prefrontal (mPF) basal ganglia (BG) circuits play an important role in integrative brain functions like movement control or cognitive/motivational behavior, respectively. Although these neuronal networks express 5-HT2A receptors, the expression is higher in associative/limbic structures than in the motor ones. We show a topographical-dependent dissociation in the effects triggered by the 5HT2A agonist TCB-2, which specifically increases the medial substantia nigra pars reticulata neuron activity and has a preferential action on mPF information processing through the striato-nigral direct pathway. These are very likely to be 5-HT2A receptor-mediated effects that require mobilization of the endogenous 5-HT system. These findings provide evidence about the specific signature of 5-HT2A receptors on the dynamic regulation of BG circuits.
Keywords: 5-HT2A receptor; TCB-2; basal ganglia circuits; fluoxetine; pCPA; serotonin.
Copyright © 2023 the authors.
Figures
Similar articles
-
Basal ganglia and processing of cortical information: functional interactions between trans-striatal and trans-subthalamic circuits in the substantia nigra pars reticulata.Neuroscience. 2003;117(4):931-8. doi: 10.1016/s0306-4522(02)00824-2. Neuroscience. 2003. PMID: 12654344
-
Cannabinoids differentially modulate cortical information transmission through the sensorimotor or medial prefrontal basal ganglia circuits.Br J Pharmacol. 2019 Apr;176(8):1156-1169. doi: 10.1111/bph.14613. Epub 2019 Mar 18. Br J Pharmacol. 2019. PMID: 30735570 Free PMC article.
-
Dopaminergic denervation impairs cortical motor and associative/limbic information processing through the basal ganglia and its modulation by the CB1 receptor.Neurobiol Dis. 2021 Jan;148:105214. doi: 10.1016/j.nbd.2020.105214. Epub 2020 Dec 3. Neurobiol Dis. 2021. PMID: 33278598
-
The connections of the primate subthalamic nucleus: indirect pathways and the open-interconnected scheme of basal ganglia-thalamocortical circuitry.Brain Res Brain Res Rev. 1997 Feb;23(1-2):62-78. doi: 10.1016/s0165-0173(96)00018-5. Brain Res Brain Res Rev. 1997. PMID: 9063587 Review.
-
Functional anatomy of the basal ganglia: limbic aspects.Rev Neurol (Paris). 2012 Aug-Sep;168(8-9):569-75. doi: 10.1016/j.neurol.2012.06.015. Epub 2012 Aug 14. Rev Neurol (Paris). 2012. PMID: 22902172 Review.
References
-
- Aghajanian GK, Foote WE, Sheard MH (1970) Action of psychotogenic drugs on single midbrain raphe neurons. J Pharmacol Exp Ther 171:178–187. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous