

Structural and dynamic basis of DNA capture and translocation by mitochondrial Twinkle helicase
- PMID: 36400570
- PMCID: PMC9723800
- DOI: 10.1093/nar/gkac1089
Structural and dynamic basis of DNA capture and translocation by mitochondrial Twinkle helicase
Abstract
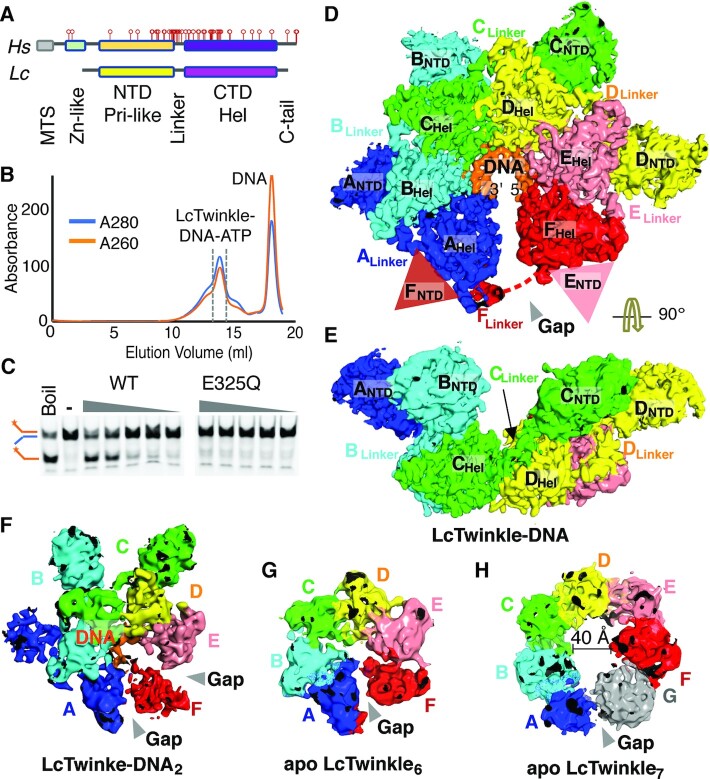
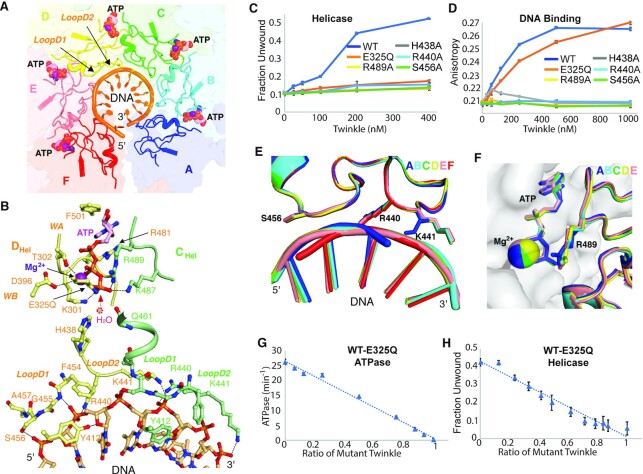
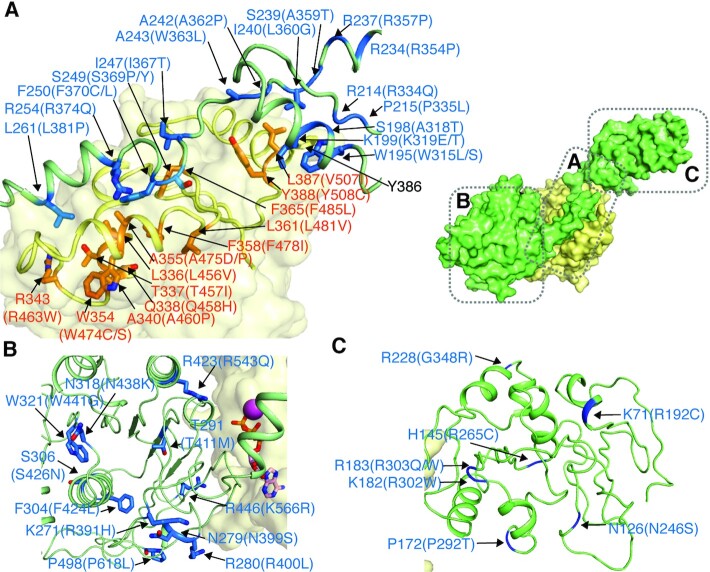
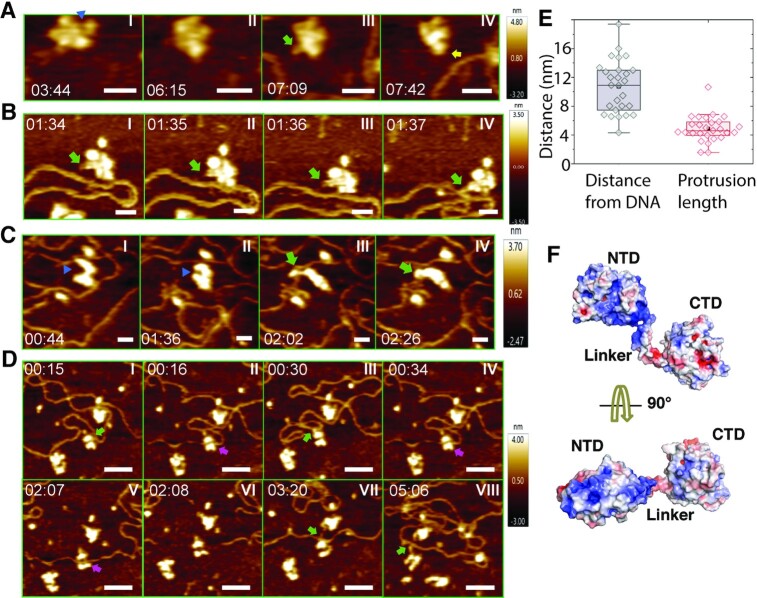
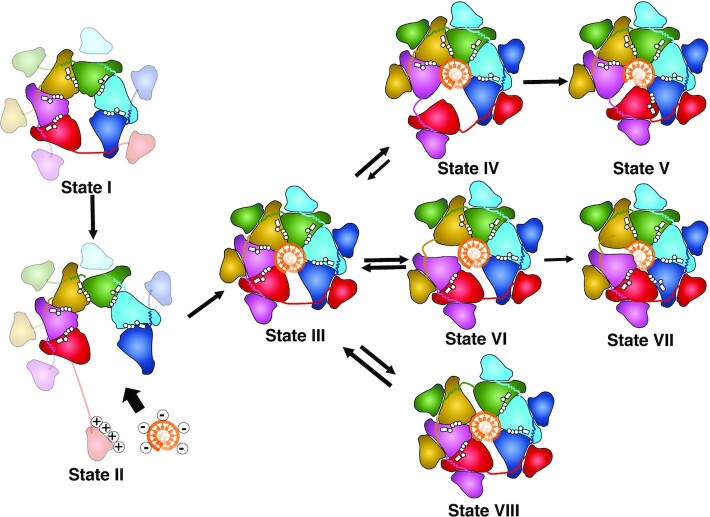
Twinkle is a mitochondrial replicative helicase which can self-load onto and unwind mitochondrial DNA. Nearly 60 mutations on Twinkle have been linked to human mitochondrial diseases. Using cryo-electron microscopy (cryo-EM) and high-speed atomic force microscopy (HS-AFM), we obtained the atomic-resolution structure of a vertebrate Twinkle homolog with DNA and captured in real-time how Twinkle is self-loaded onto DNA. Our data highlight the important role of the non-catalytic N-terminal domain of Twinkle. The N-terminal domain directly contacts the C-terminal helicase domain, and the contact interface is a hotspot for disease-related mutations. Mutations at the interface destabilize Twinkle hexamer and reduce helicase activity. With HS-AFM, we observed that a highly dynamic Twinkle domain, which is likely to be the N-terminal domain, can protrude ∼5 nm to transiently capture nearby DNA and initialize Twinkle loading onto DNA. Moreover, structural analysis and subunit doping experiments suggest that Twinkle hydrolyzes ATP stochastically, which is distinct from related helicases from bacteriophages.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Method for the structural analysis of Twinkle mitochondrial DNA helicase by cryo-EM.Methods. 2022 Sep;205:263-270. doi: 10.1016/j.ymeth.2022.06.012. Epub 2022 Jun 30. Methods. 2022. PMID: 35779765 Free PMC article.
-
The N-terminal domain of human mitochondrial helicase Twinkle has DNA-binding activity crucial for supporting processive DNA synthesis by polymerase γ.J Biol Chem. 2023 Jan;299(1):102797. doi: 10.1016/j.jbc.2022.102797. Epub 2022 Dec 14. J Biol Chem. 2023. PMID: 36528058 Free PMC article.
-
Single-molecule level structural dynamics of DNA unwinding by human mitochondrial Twinkle helicase.J Biol Chem. 2020 Apr 24;295(17):5564-5576. doi: 10.1074/jbc.RA120.012795. Epub 2020 Mar 25. J Biol Chem. 2020. PMID: 32213598 Free PMC article.
-
TWINKLE and Other Human Mitochondrial DNA Helicases: Structure, Function and Disease.Genes (Basel). 2020 Apr 9;11(4):408. doi: 10.3390/genes11040408. Genes (Basel). 2020. PMID: 32283748 Free PMC article. Review.
-
Borrowing nuclear DNA helicases to protect mitochondrial DNA.Int J Mol Sci. 2015 May 13;16(5):10870-87. doi: 10.3390/ijms160510870. Int J Mol Sci. 2015. PMID: 25984607 Free PMC article. Review.
Cited by
-
Autoregulation of the real-time kinetics of the human mitochondrial replicative helicase.Nat Commun. 2025 Jul 1;16(1):5460. doi: 10.1038/s41467-025-60289-0. Nat Commun. 2025. PMID: 40592829 Free PMC article.
-
A Bacteriophage-Derived Primase-Helicase Orchestrates Plant Organellar DNA Replication.Physiol Plant. 2025 Jul-Aug;177(4):e70379. doi: 10.1111/ppl.70379. Physiol Plant. 2025. PMID: 40620047 Free PMC article.
-
Twinkle-Catalyzed Toehold-Mediated DNA Strand Displacement Reaction.J Am Chem Soc. 2023 Nov 2:10.1021/jacs.3c04970. doi: 10.1021/jacs.3c04970. Online ahead of print. J Am Chem Soc. 2023. PMID: 37917930 Free PMC article.
-
Structural basis for the concerted antiphage activity in the SIR2-HerA system.Nucleic Acids Res. 2024 Oct 14;52(18):11336-11348. doi: 10.1093/nar/gkae750. Nucleic Acids Res. 2024. PMID: 39217465 Free PMC article.
-
Single-molecule imaging of genome maintenance proteins encountering specific DNA sequences and structures.DNA Repair (Amst). 2023 Aug;128:103528. doi: 10.1016/j.dnarep.2023.103528. Epub 2023 Jun 24. DNA Repair (Amst). 2023. PMID: 37392578 Free PMC article. Review.
References
-
- Gustafsson C.M., Falkenberg M., Larsson N.G.. Maintenance and expression of mammalian mitochondrial DNA. Annu. Rev. Biochem. 2016; 85:133–160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
