

Structural and functional asymmetry of RING trimerization controls priming and extension events in TRIM5α autoubiquitylation
- PMID: 36402777
- PMCID: PMC9675739
- DOI: 10.1038/s41467-022-34920-3
Structural and functional asymmetry of RING trimerization controls priming and extension events in TRIM5α autoubiquitylation
Abstract
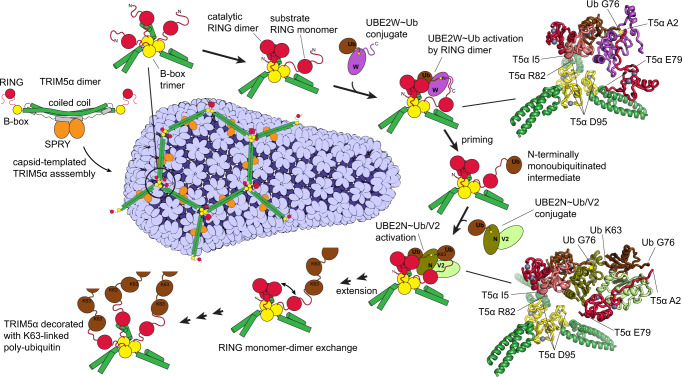
TRIM5α is an E3 ubiquitin ligase of the TRIM family that binds to the capsids of primate immunodeficiency viruses and blocks viral replication after cell entry. Here we investigate how synthesis of K63-linked polyubiquitin is upregulated by transient proximity of three RING domains in honeycomb-like assemblies formed by TRIM5α on the surface of the retroviral capsid. Proximity of three RINGs creates an asymmetric arrangement, in which two RINGs form a catalytic dimer that activates E2-ubiquitin conjugates and the disordered N-terminus of the third RING acts as the substrate for N-terminal autoubiquitylation. RING dimerization is required for activation of the E2s that contribute to the antiviral function of TRIM5α, UBE2W and heterodimeric UBE2N/V2, whereas the proximity of the third RING enhances the rate of each of the two distinct steps in the autoubiquitylation process: the initial N-terminal monoubiquitylation (priming) of TRIM5α by UBE2W and the subsequent extension of the K63-linked polyubiquitin chain by UBE2N/V2. The mechanism we describe explains how recognition of infection-associated epitope patterns by TRIM proteins initiates polyubiquitin-mediated downstream events in innate immunity.
© 2022. The Author(s).
Conflict of interest statement
A pending patent application PCT/US2022/048268 lists the Board of Regents, The University of Texas System as the applicant and F.H. and D.N.I. as inventors. D.N.I. is a co-founder and a shareholder of E3 Biosciences LLC, a commercial entity that manufactures reagents described in this study for use in academic and commercial research. The remaining authors declare no other competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
