

Biofabrication of synthetic human liver tissue with advanced programmable functions
- PMID: 36404924
- PMCID: PMC9672940
- DOI: 10.1016/j.isci.2022.105503
Biofabrication of synthetic human liver tissue with advanced programmable functions
Abstract
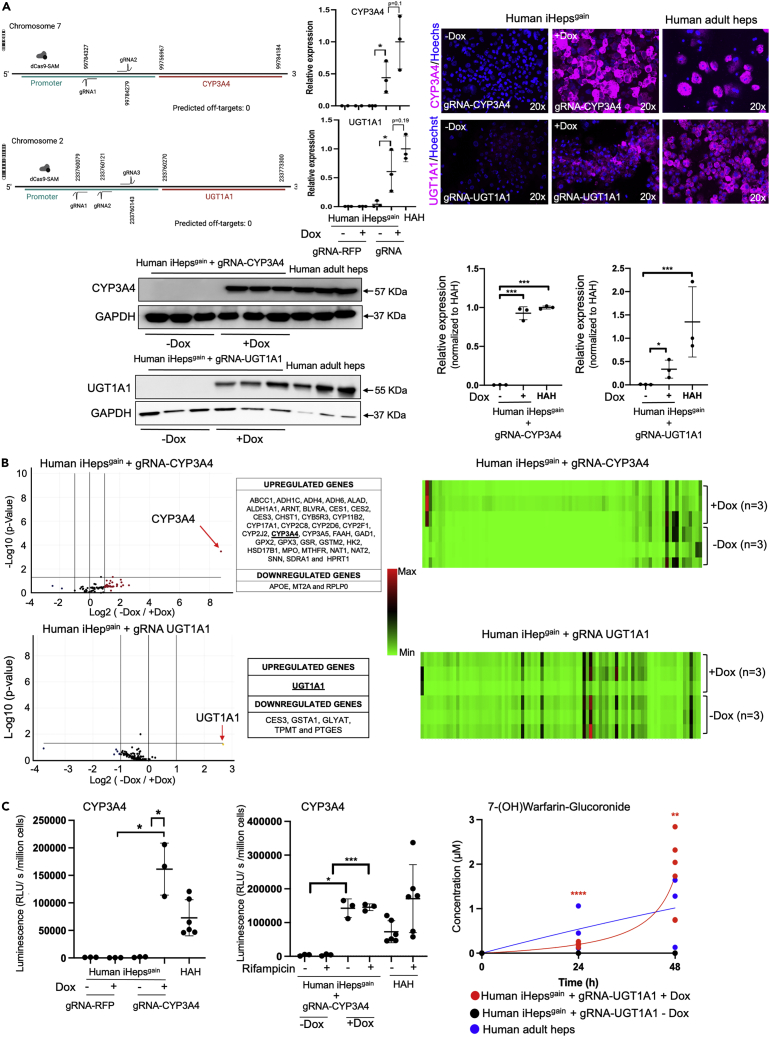
Advances in cellular engineering, as well as gene, and cell therapy, may be used to produce human tissues with programmable genetically enhanced functions designed to model and/or treat specific diseases. Fabrication of synthetic human liver tissue with these programmable functions has not been described. By generating human iPSCs with target gene expression controlled by a guide RNA-directed CRISPR-Cas9 synergistic-activation-mediator, we produced synthetic human liver tissues with programmable functions. Such iPSCs were guide-RNA-treated to enhance expression of the clinically relevant CYP3A4 and UGT1A1 genes, and after hepatocyte-directed differentiation, cells demonstrated enhanced functions compared to those found in primary human hepatocytes. We then generated human liver tissue with these synthetic human iPSC-derived hepatocytes (iHeps) and other non-parenchymal cells demonstrating advanced programmable functions. Fabrication of synthetic human liver tissue with modifiable functional genetic programs may be a useful tool for drug discovery, investigating biology, and potentially creating bioengineered organs with specialized functions.
Keywords: Bioengineering; Biological sciences; Tissue engineering.
© 2022 The Authors.
Conflict of interest statement
A.C.-H., K.T., I.J.F., and A.S.-G. are inventors on a provisional international patent application that describes hepatic differentiation of human pluripotent stem cells and liver repopulation (PCT/US2018/018032) and a provisional international patent application that describes the use of human-induced pluripotent stem cells for highly genetic engineering (PCT/US2017/044719). A.S.-G., J.G.-L., A.C.-H., and I.J.F. are co-founders and have a financial interest in Von Baer Wolff, Inc. a company focused on biofabrication of autologous human hepatocytes from stem cells technology. I.J.F. and A.S.-G. are co-founders and have a financial interest in Pittsburgh ReLiver Inc, a company focused on programming liver failure and their interests are managed by the Conflict-of-Interest Office at the University of Pittsburgh in accordance with their policies.
Figures




References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
