

Uncoupling cell division and cytokinesis during germline development in metazoans
- PMID: 36407108
- PMCID: PMC9669650
- DOI: 10.3389/fcell.2022.1001689
Uncoupling cell division and cytokinesis during germline development in metazoans
Abstract
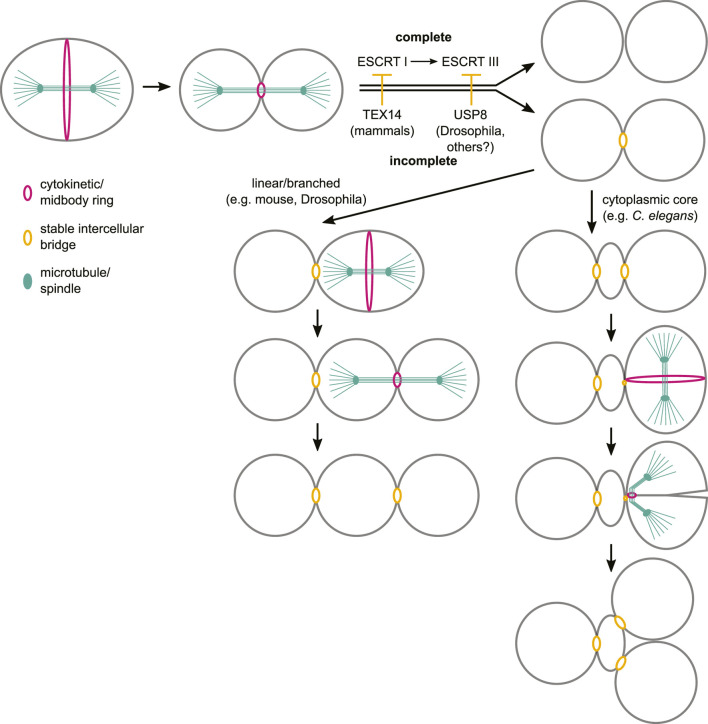
The canonical eukaryotic cell cycle ends with cytokinesis, which physically divides the mother cell in two and allows the cycle to resume in the newly individualized daughter cells. However, during germline development in nearly all metazoans, dividing germ cells undergo incomplete cytokinesis and germ cells stay connected by intercellular bridges which allow the exchange of cytoplasm and organelles between cells. The near ubiquity of incomplete cytokinesis in animal germ lines suggests that this is an ancient feature that is fundamental for the development and function of this tissue. While cytokinesis has been studied for several decades, the mechanisms that enable regulated incomplete cytokinesis in germ cells are only beginning to emerge. Here we review the current knowledge on the regulation of germ cell intercellular bridge formation, focusing on findings made using mouse, Drosophila melanogaster and Caenorhabditis elegans as experimental systems.
Keywords: germ cells; germline development; incomplete cytokinesis; intercellular bridges; metazoan.
Copyright © 2022 Gerhold, Labbé and Singh.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




Similar articles
-
The initial expansion of the C. elegans syncytial germ line is coupled to incomplete primordial germ cell cytokinesis.Development. 2021 Sep 15;148(18):dev199633. doi: 10.1242/dev.199633. Epub 2021 Jul 21. Development. 2021. PMID: 34195824 Free PMC article.
-
From primordial germ cells to spermatids in Caenorhabditis elegans.Semin Cell Dev Biol. 2022 Jul;127:110-120. doi: 10.1016/j.semcdb.2021.12.005. Epub 2021 Dec 17. Semin Cell Dev Biol. 2022. PMID: 34930663 Review.
-
A Sterile 20 Family Kinase and Its Co-factor CCM-3 Regulate Contractile Ring Proteins on Germline Intercellular Bridges.Curr Biol. 2017 Mar 20;27(6):860-867. doi: 10.1016/j.cub.2017.01.058. Epub 2017 Mar 9. Curr Biol. 2017. PMID: 28285996 Free PMC article.
-
C. elegans Anillin proteins regulate intercellular bridge stability and germline syncytial organization.J Cell Biol. 2014 Jul 7;206(1):129-43. doi: 10.1083/jcb.201310117. Epub 2014 Jun 30. J Cell Biol. 2014. PMID: 24982432 Free PMC article.
-
Germ cell intercellular bridges.Cold Spring Harb Perspect Biol. 2011 Aug 1;3(8):a005850. doi: 10.1101/cshperspect.a005850. Cold Spring Harb Perspect Biol. 2011. PMID: 21669984 Free PMC article. Review.
Cited by
-
Examination of germline and somatic intercellular bridges in Hydra vulgaris reveals insights into the evolutionarily conserved mechanism of intercellular bridge formation.bioRxiv [Preprint]. 2025 Feb 20:2025.02.19.639158. doi: 10.1101/2025.02.19.639158. bioRxiv. 2025. PMID: 40027827 Free PMC article. Preprint.
-
ARF6, a component of intercellular bridges, is essential for spermatogenesis in mice.Dev Biol. 2024 Apr;508:46-63. doi: 10.1016/j.ydbio.2024.01.007. Epub 2024 Jan 17. Dev Biol. 2024. PMID: 38242343 Free PMC article.
-
Testis exposure to unopposed/elevated activin A in utero affects somatic and germ cells and alters steroid levels mimicking phthalate exposure.Front Endocrinol (Lausanne). 2023 Sep 1;14:1234712. doi: 10.3389/fendo.2023.1234712. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37727456 Free PMC article.
-
A key role for centralspindlin and Ect2 in the development of multicellularity and the emergence of Metazoa.Curr Biol. 2025 Jul 7;35(13):3251-3262.e1. doi: 10.1016/j.cub.2025.05.052. Epub 2025 Jun 17. Curr Biol. 2025. PMID: 40532703
-
Non-Mendelian transmission of X chromosomes: mechanisms and impact on sex ratios and population dynamics in different breeding systems.Biochem Soc Trans. 2024 Aug 28;52(4):1777-1784. doi: 10.1042/BST20231411. Biochem Soc Trans. 2024. PMID: 39149984 Free PMC article. Review.
References
-
- Anderson E., Huebner E. (1968). Development of the oocyte and its accessory cells of the polychaete, Diopatra cuprea (Bosc). J. Morphol. 126, 163–197. 10.1002/jmor.1051260203 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases