

The ESX-1 Substrate PPE68 Has a Key Function in ESX-1-Mediated Secretion in Mycobacterium marinum
- PMID: 36409073
- PMCID: PMC9765416
- DOI: 10.1128/mbio.02819-22
The ESX-1 Substrate PPE68 Has a Key Function in ESX-1-Mediated Secretion in Mycobacterium marinum
Abstract
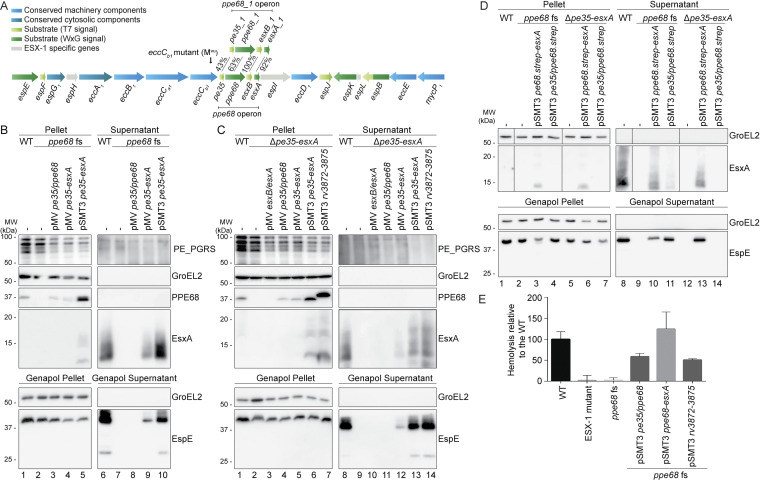
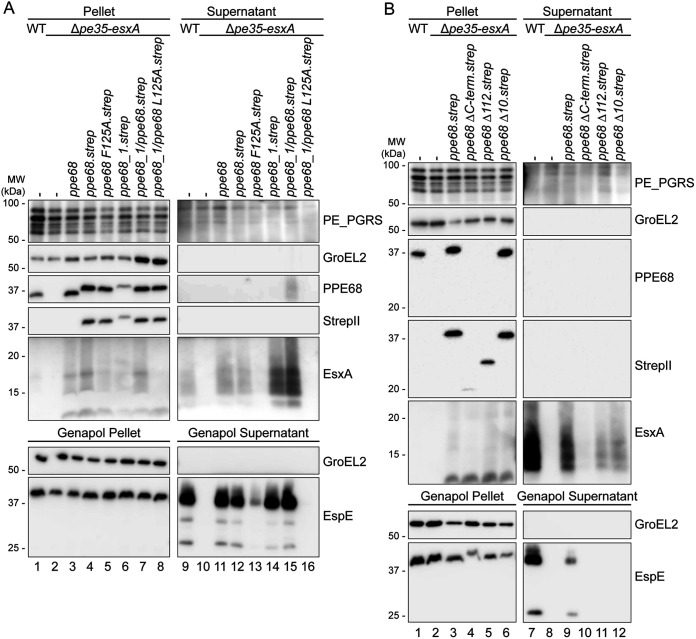
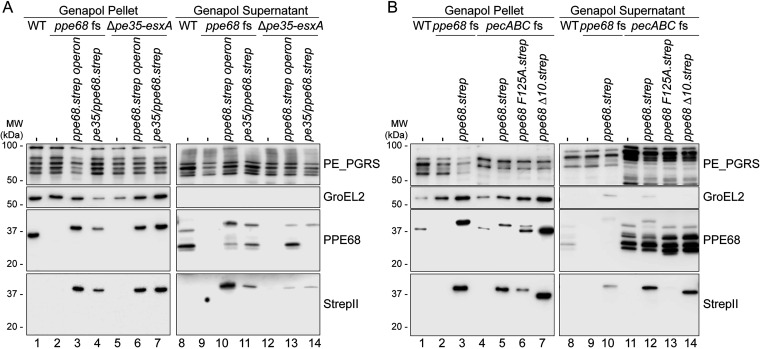
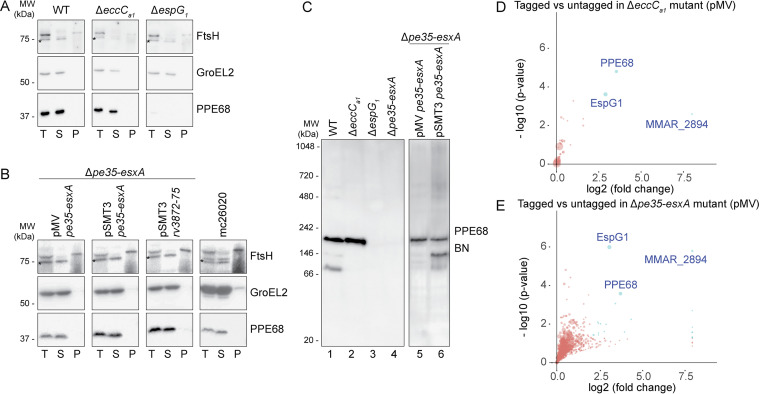
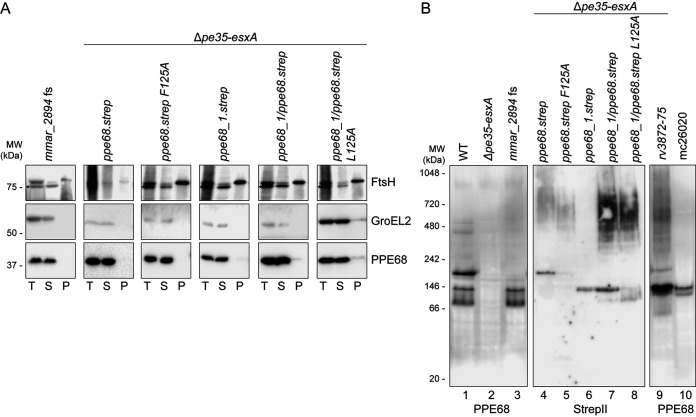
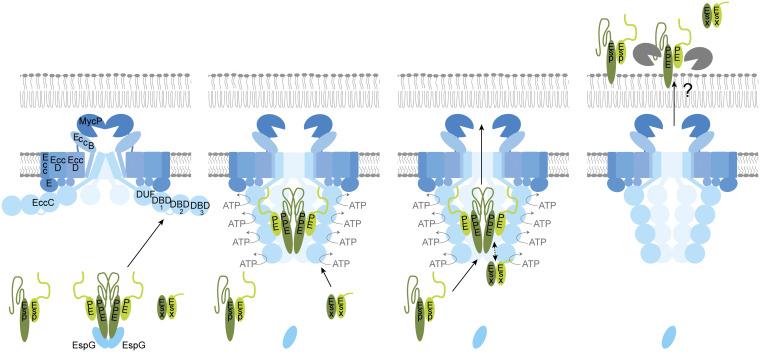
Mycobacteria use specialized type VII secretion systems (T7SSs) to secrete proteins across their diderm cell envelope. One of the T7SS subtypes, named ESX-1, is a major virulence determinant in pathogenic species such as Mycobacterium tuberculosis and the fish pathogen Mycobacterium marinum. ESX-1 secretes a variety of substrates, called Esx, PE, PPE, and Esp proteins, at least some of which are folded heterodimers. Investigation into the functions of these substrates is problematic, because of the intricate network of codependent secretion between several ESX-1 substrates. Here, we describe the ESX-1 substrate PPE68 as essential for secretion of the highly immunogenic substrates EsxA and EspE via the ESX-1 system in M. marinum. While secreted PPE68 is processed on the cell surface, the majority of cell-associated PPE68 of M. marinum and M. tuberculosis is present in a cytosolic complex with its PE partner and the EspG1 chaperone. Interfering with the binding of EspG1 to PPE68 blocked its export and the secretion of EsxA and EspE. In contrast, esxA was not required for the secretion of PPE68, revealing a hierarchy in codependent secretion. Remarkably, the final 10 residues of PPE68, a negatively charged domain, seem essential for EspE secretion, but not for the secretion of EsxA and of PPE68 itself. This indicates that distinctive domains of PPE68 are involved in secretion of the different ESX-1 substrates. Based on these findings, we propose a mechanistic model for the central role of PPE68 in ESX-1-mediated secretion and substrate codependence. IMPORTANCE Pathogenic mycobacteria, such Mycobacterium tuberculosis and Mycobacterium marinum, use a type VII secretion system (T7SS) subtype, called ESX-1, to mediate intracellular survival via phagosomal rupture and subsequent translocation of the mycobacterium to the host cytosol. Identifying the ESX-1 substrate that is responsible for this process is problematic because of the intricate network of codependent secretion between ESX-1 substrates. Here, we show the central role of the ESX-1 substrate PPE68 for the secretion of ESX-1 substrates in Mycobacterium marinum. Unravelling the mechanism of codependent secretion will aid the functional understanding of T7SSs and will allow the analysis of the individual roles of ESX-1 substrates in the virulence caused by the significant human pathogen Mycobacterium tuberculosis.
Keywords: ESX-1; EsxA; Mycobacterium; PPE; chaperones; protein transport; protein-protein interactions; tuberculosis; type VII secretion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Houben D, Demangel C, van Ingen J, Perez J, Baldeon L, Abdallah AM, Caleechurn L, Bottai D, van Zon M, de Punder K, van der Laan T, Kant A, Bossers-de Vries R, Willemsen P, Bitter W, van Soolingen D, Brosch R, van der Wel N, Peters PJ. 2012. ESX-1-mediated translocation to the cytosol controls virulence of mycobacteria. Cell Microbiol 14:1287–1298. doi: 10.1111/j.1462-5822.2012.01799.x. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
