

The RNA-Binding Protein ProQ Promotes Antibiotic Persistence in Salmonella
- PMID: 36409088
- PMCID: PMC9765298
- DOI: 10.1128/mbio.02891-22
The RNA-Binding Protein ProQ Promotes Antibiotic Persistence in Salmonella
Abstract
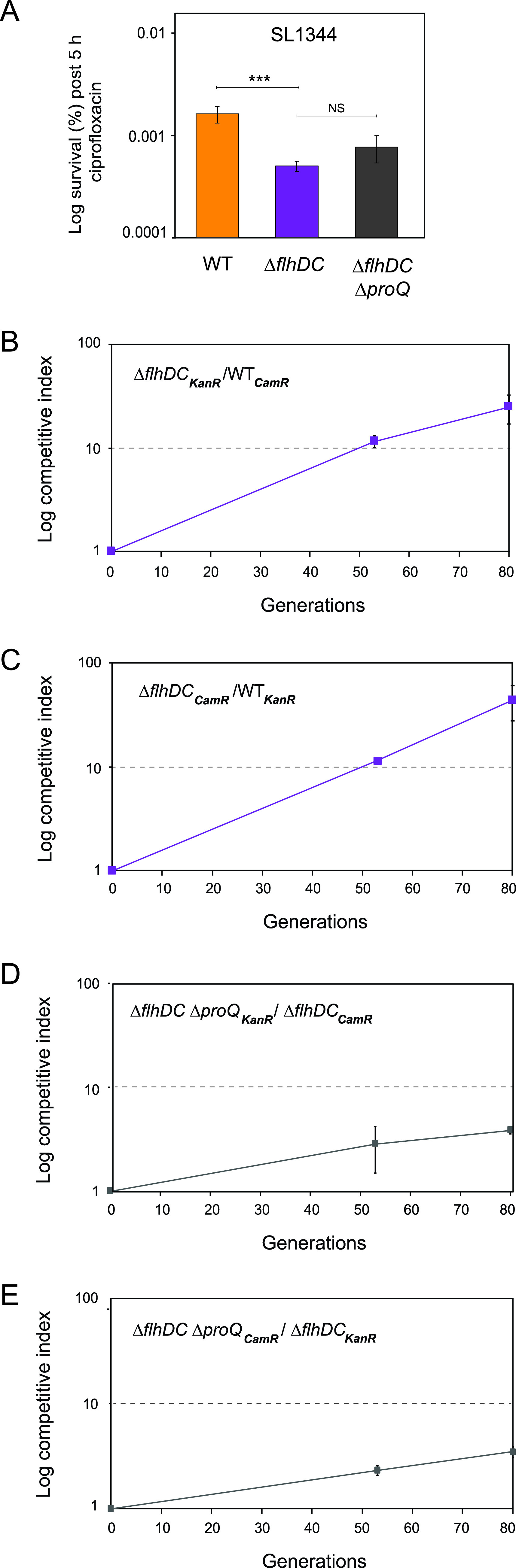
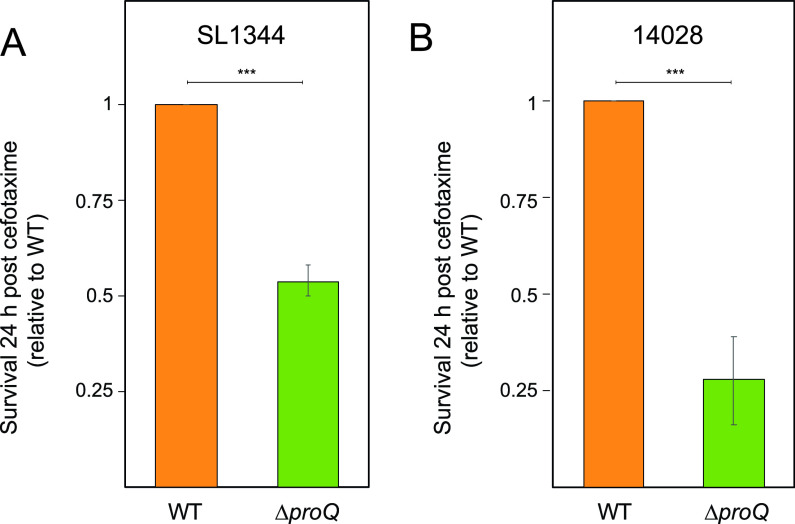
Bacterial populations can survive exposure to antibiotics through transient phenotypic and gene expression changes. These changes can be attributed to a small subpopulation of bacteria, giving rise to antibiotic persistence. Although this phenomenon has been known for decades, much remains to be learned about the mechanisms that drive persister formation. The RNA-binding protein ProQ has recently emerged as a global regulator of gene expression. Here, we show that ProQ impacts persister formation in Salmonella. In vitro, ProQ contributes to growth arrest in a subset of cells that are able to survive treatment at high concentrations of different antibiotics. The underlying mechanism for ProQ-dependent persister formation involves the activation of metabolically costly processes, including the flagellar pathway and the type III protein secretion system encoded on Salmonella pathogenicity island 2. Importantly, we show that the ProQ-dependent phenotype is relevant during macrophage infection and allows Salmonella to survive the combined action of host immune defenses and antibiotics. Together, our data highlight the importance of ProQ in Salmonella persistence and pathogenesis. IMPORTANCE Bacteria can avoid eradication by antibiotics through a phenomenon known as persistence. Persister cells arise through phenotypic heterogeneity and constitute a small fraction of dormant cells within a population of actively growing bacteria, which is susceptible to antibiotic killing. In this study, we show that ProQ, an RNA-binding protein and global regulator of gene expression, promotes persisters in the human pathogen Salmonella enterica serovar Typhimurium. Bacteria lacking the proQ gene outcompete wild-type bacteria under laboratory conditions, are less prone to enter growth dormancy, and form fewer persister cells. The basis for these phenotypes lies in ProQ's ability to activate energy-consuming cellular processes, including flagellar motility and protein secretion. Importantly, we show that ProQ contributes to the persister phenotype during Salmonella infection of macrophages, indicating an important role of this global regulator in Salmonella pathogenesis.
Keywords: ProQ; RNA-binding protein; Salmonella; antibiotic persistence; antibiotic persisters; flagella; flagellar gene regulation; persister formation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
The Major RNA-Binding Protein ProQ Impacts Virulence Gene Expression in Salmonella enterica Serovar Typhimurium.mBio. 2019 Jan 2;10(1):e02504-18. doi: 10.1128/mBio.02504-18. mBio. 2019. PMID: 30602583 Free PMC article.
-
ProQ-dependent activation of Salmonella virulence genes mediated by post-transcriptional control of PhoP synthesis.mSphere. 2024 Mar 26;9(3):e0001824. doi: 10.1128/msphere.00018-24. Epub 2024 Feb 27. mSphere. 2024. PMID: 38411119 Free PMC article.
-
Prophage Gifsy-1 Induction in Salmonella enterica Serovar Typhimurium Reduces Persister Cell Formation after Ciprofloxacin Exposure.Microbiol Spectr. 2023 Aug 17;11(4):e0187423. doi: 10.1128/spectrum.01874-23. Epub 2023 Jun 12. Microbiol Spectr. 2023. PMID: 37306609 Free PMC article.
-
Studying antibiotic persistence in vivo using the model organism Salmonella Typhimurium.Curr Opin Microbiol. 2022 Dec;70:102224. doi: 10.1016/j.mib.2022.102224. Epub 2022 Nov 3. Curr Opin Microbiol. 2022. PMID: 36335713 Review.
-
Toxin-Antitoxin Systems: A Key Role on Persister Formation in Salmonella enterica Serovar Typhimurium.Infect Drug Resist. 2022 Oct 3;15:5813-5829. doi: 10.2147/IDR.S378157. eCollection 2022. Infect Drug Resist. 2022. PMID: 36213766 Free PMC article. Review.
Cited by
-
RNA recognition by minimal ProQ from Neisseria meningitidis.RNA. 2025 Mar 18;31(4):549-565. doi: 10.1261/rna.080207.124. RNA. 2025. PMID: 39875175 Free PMC article.
-
ProQ prevents mRNA degradation through inhibition of poly(A) polymerase.Nucleic Acids Res. 2025 Feb 27;53(5):gkaf103. doi: 10.1093/nar/gkaf103. Nucleic Acids Res. 2025. PMID: 40036335 Free PMC article.
-
Bacterial persisters: molecular mechanisms and therapeutic development.Signal Transduct Target Ther. 2024 Jul 17;9(1):174. doi: 10.1038/s41392-024-01866-5. Signal Transduct Target Ther. 2024. PMID: 39013893 Free PMC article. Review.
-
Disruption of sulfur transferase complex increases bacterial intramacrophage persistence.PLoS Pathog. 2025 May 14;21(5):e1013136. doi: 10.1371/journal.ppat.1013136. eCollection 2025 May. PLoS Pathog. 2025. PMID: 40367211 Free PMC article.
-
The FinO/ProQ-like protein PA2582 impacts antimicrobial resistance in Pseudomonas aeruginosa.Front Microbiol. 2024 Jun 26;15:1422742. doi: 10.3389/fmicb.2024.1422742. eCollection 2024. Front Microbiol. 2024. PMID: 39011145 Free PMC article.
References
-
- Balaban NQ, Helaine S, Lewis K, Ackermann M, Aldridge B, Andersson DI, Brynildsen MP, Bumann D, Camilli A, Collins JJ, Dehio C, Fortune S, Ghigo JM, Hardt WD, Harms A, Heinemann M, Hung DT, Jenal U, Levin BR, Michiels J, Storz G, Tan MW, Tenson T, Van Melderen L, Zinkernagel A. 2019. Definitions and guidelines for research on antibiotic persistence. Nat Rev Microbiol 17:441–448. doi:10.1038/s41579-019-0196-3. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
