

Oxidative stress-induced FAK activation contributes to uterine serous carcinoma aggressiveness
- PMID: 36409196
- PMCID: PMC9812840
- DOI: 10.1002/1878-0261.13346
Oxidative stress-induced FAK activation contributes to uterine serous carcinoma aggressiveness
Abstract
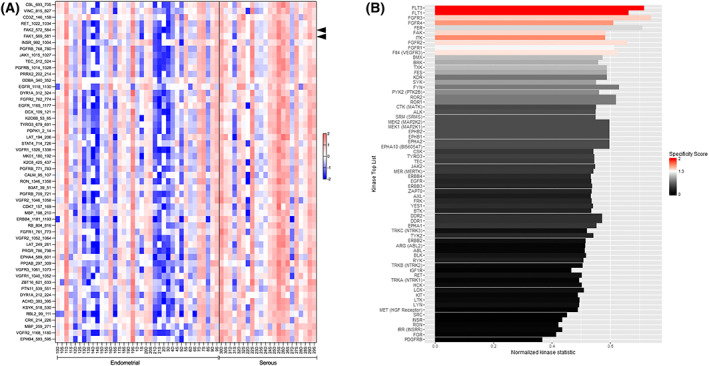
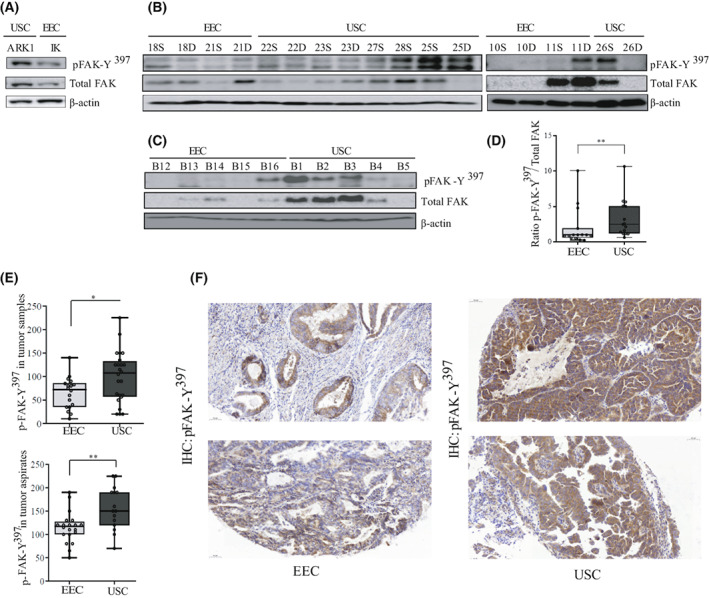
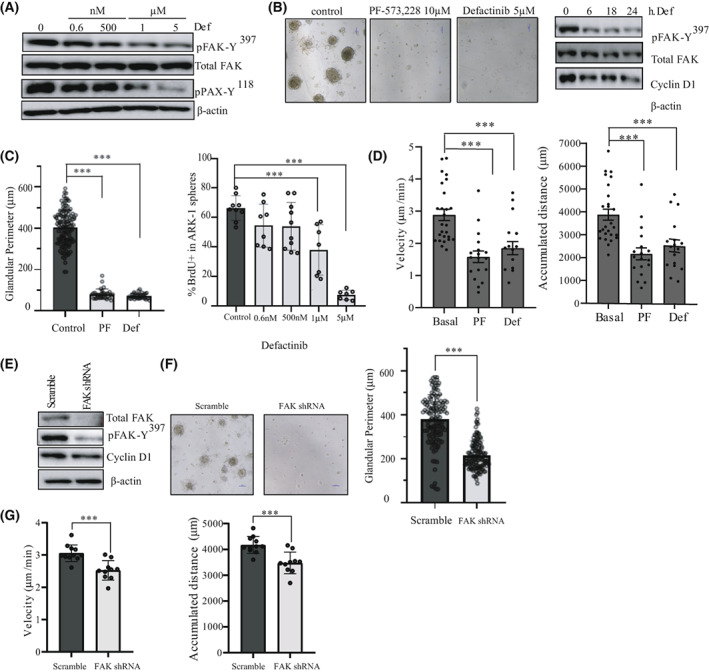
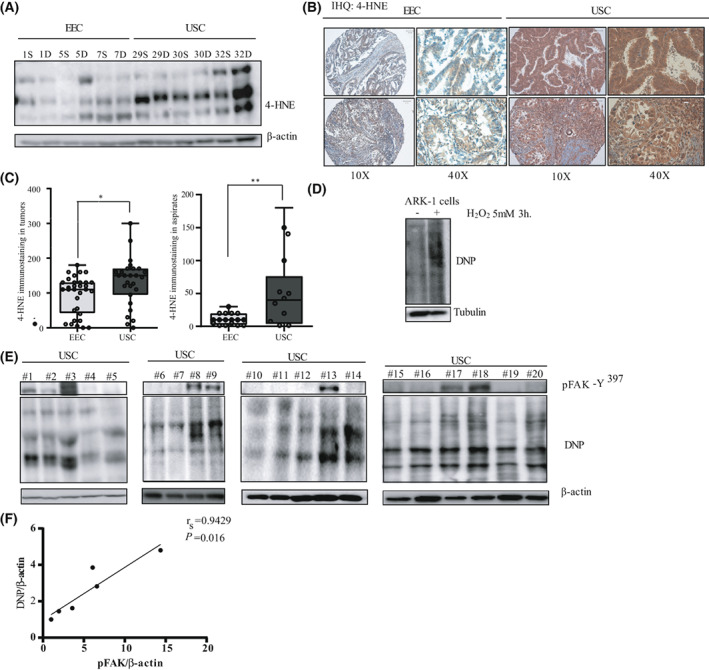
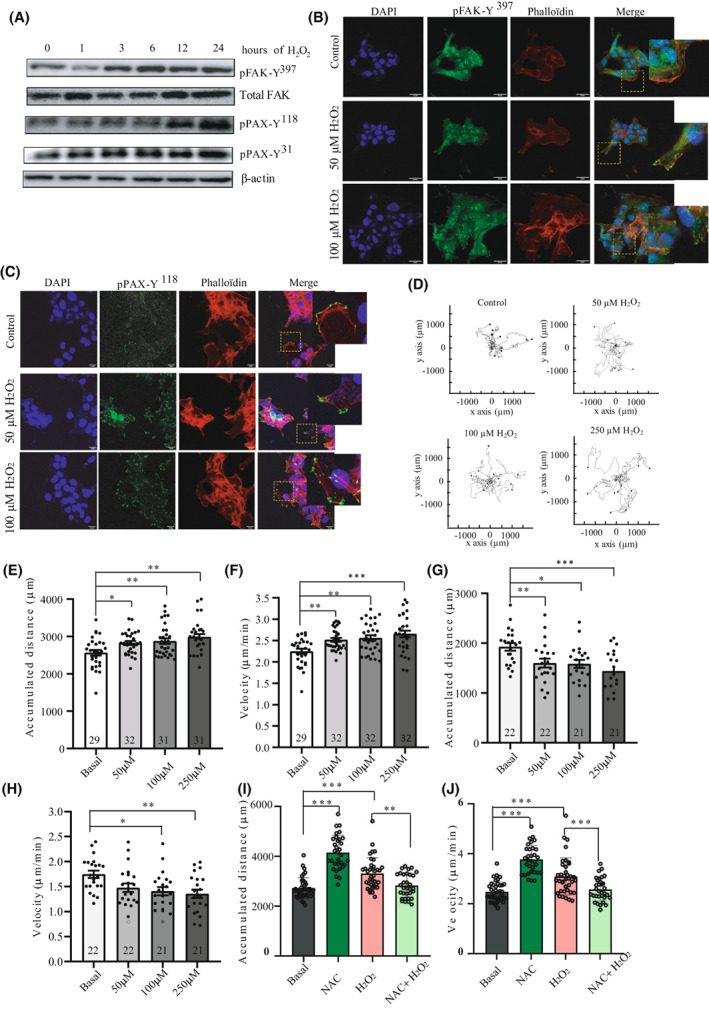
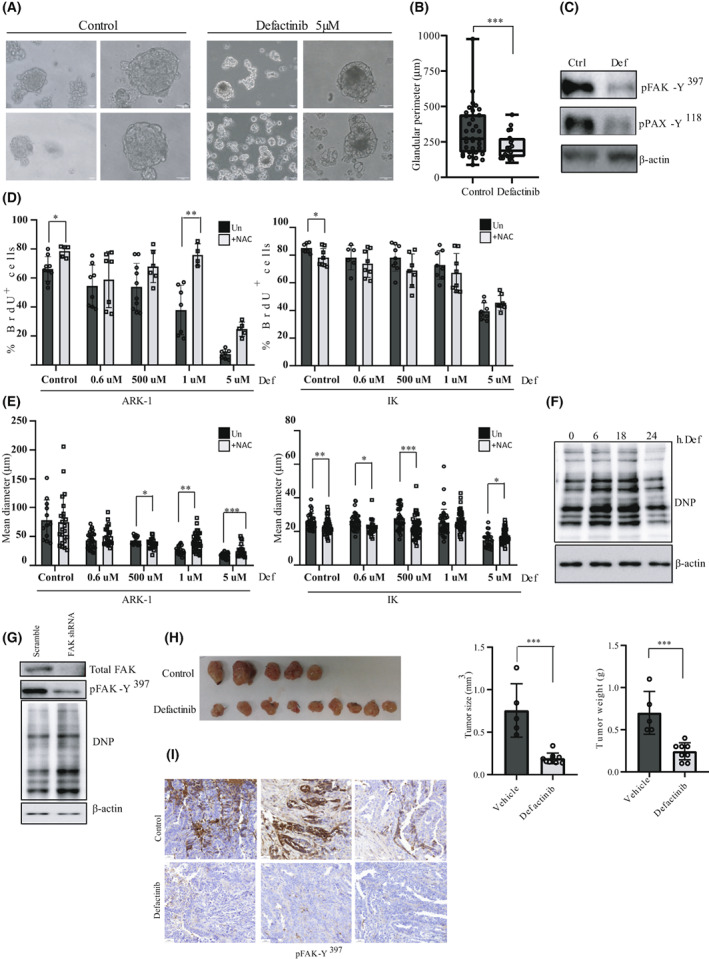
Uterine serous carcinoma (USC) is an aggressive form of endometrial cancer (EC), characterized by its high propensity for metastases. In fact, while endometrioid endometrial carcinoma (EEC), which accounts for 85% of EC, presents a good prognosis, USC is the most frequently fatal. Herein, we used for the first time a peptide-based tyrosine-kinase-activity profiling approach to quantify the changes in tyrosine kinase activation between USC and EEC. Among the tyrosine kinases highly activated in USC, we identified focal adhesion kinase (FAK). We conducted mechanistic studies using cellular models. In a USC cell line, targeting FAK either by inhibitors PF-573228 and defactinib (VS-6063) or by gene silencing limits 3D cell growth and reduces cell migration. Moreover, results from our studies suggest that oxidative stress is increased in USC tumors compared to EEC ones. Reactive oxygen species (ROS) induce tyrosine phosphorylation of FAK and a concomitant tyrosine phosphorylation of paxillin, a mediator of FAK signal transduction. Mechanistically, by tracking hundreds of individual cells per condition, we show that ROS increased cell distance and migration velocity, highlighting the role of ROS-FAK-PAX signaling in cell migration. Both defactinib and ROS scavenger N-acetylcysteine (NAC) revert this effect, pointing toward ROS as potential culprits for the increase in USC cell motility. A proof of concept of the role of FAK in controlling cell growth was obtained in in vivo experiments using cancer-tissue-originated spheroids (CTOS) and a patient-derived orthotopic xenograft model (orthoxenograft/PDOX). Defactinib reduces cell proliferation and protein oxidation, supporting a pro-tumoral antioxidant role of FAK, whereas antioxidant NAC reverts FAK inhibitor effects. Overall, our data points to ROS-mediated FAK activation in USC as being responsible for the poor prognosis of this tumor type and emphasize the potential of FAK inhibition for USC treatment.
Keywords: focal adhesion kinase; migration; reactive oxygen species; uterine serous carcinoma.
© 2022 The Authors. Molecular Oncology published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
A. Villanueva and A. Vidal are co‐founders of Xenopat S.L. The rest of authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
