

Exercise increases information content and affects long-term stability of hippocampal place codes
- PMID: 36417871
- PMCID: PMC9715913
- DOI: 10.1016/j.celrep.2022.111695
Exercise increases information content and affects long-term stability of hippocampal place codes
Abstract
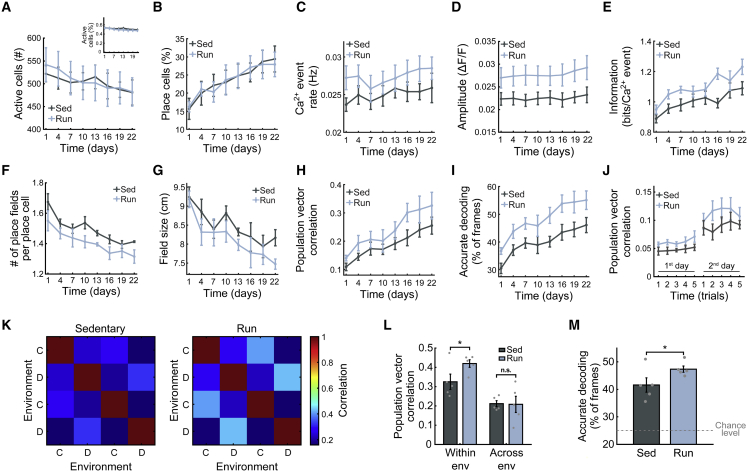
Physical exercise is known to augment brain functioning, improving memory and cognition. However, while some of the physiological effects of physical activity on the brain are known, little is known about its effects on the neural code. Using calcium imaging in freely behaving mice, we study how voluntary exercise affects the quality and long-term stability of hippocampal place codes. We find that running accelerates the emergence of a more informative spatial code in novel environments and increases code stability over days and weeks. Paradoxically, although runners demonstrated an overall more stable place code than their sedentary peers, their place code changed faster when controlling for code quality level. A model-based simulation shows that the combination of improved code quality and faster representational drift in runners, but neither of these effects alone, could account for our results. Thus, exercise may enhance hippocampal function via a more informative and dynamic place code.
Keywords: CP: Neuroscience; adult neurogenesis; calcium imaging; hippocampus; memory; neural coding; physical activity; place cells; representational drift; running.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




Similar articles
-
Age-specific effects of voluntary exercise on memory and the older brain.Biol Psychiatry. 2013 Mar 1;73(5):435-42. doi: 10.1016/j.biopsych.2012.05.034. Epub 2012 Jul 15. Biol Psychiatry. 2013. PMID: 22795967 Free PMC article.
-
Voluntary exercise does not ameliorate spatial learning and memory deficits induced by chronic administration of nandrolone decanoate in rats.Horm Behav. 2013 Jan;63(1):158-65. doi: 10.1016/j.yhbeh.2012.10.003. Epub 2012 Oct 12. Horm Behav. 2013. PMID: 23068768
-
Organization of hippocampal CA3 into correlated cell assemblies supports a stable spatial code.Cell Rep. 2023 Feb 28;42(2):112119. doi: 10.1016/j.celrep.2023.112119. Epub 2023 Feb 17. Cell Rep. 2023. PMID: 36807137 Free PMC article.
-
[Mild Exercise Results in Robust Brain Activation and Increased Memory Function].Brain Nerve. 2018 Jul;70(7):745-752. doi: 10.11477/mf.1416201075. Brain Nerve. 2018. PMID: 29997270 Review. Japanese.
-
Exercise effects on brain and behavior in healthy mice, Alzheimer's disease and Parkinson's disease model-A systematic review and meta-analysis.Behav Brain Res. 2020 Apr 6;383:112488. doi: 10.1016/j.bbr.2020.112488. Epub 2020 Jan 25. Behav Brain Res. 2020. PMID: 31991178
Cited by
-
Adult neurogenesis improves spatial information encoding in the mouse hippocampus.Nat Commun. 2024 Jul 30;15(1):6410. doi: 10.1038/s41467-024-50699-x. Nat Commun. 2024. PMID: 39080283 Free PMC article.
-
Adult-born granule cells facilitate remapping of spatial and non-spatial representations in the dentate gyrus.Neuron. 2023 Dec 20;111(24):4024-4039.e7. doi: 10.1016/j.neuron.2023.09.016. Epub 2023 Oct 10. Neuron. 2023. PMID: 37820723 Free PMC article.
-
Exploring the interplay between running exercises, microbial diversity, and tryptophan metabolism along the microbiota-gut-brain axis.Front Microbiol. 2024 Jan 22;15:1326584. doi: 10.3389/fmicb.2024.1326584. eCollection 2024. Front Microbiol. 2024. PMID: 38318337 Free PMC article.
-
Hippocampal representations drift in stable multisensory environments.Nature. 2025 Sep;645(8080):457-465. doi: 10.1038/s41586-025-09245-y. Epub 2025 Jul 23. Nature. 2025. PMID: 40702176
-
Drift of neural ensembles driven by slow fluctuations of intrinsic excitability.Elife. 2024 May 7;12:RP88053. doi: 10.7554/eLife.88053. Elife. 2024. PMID: 38712831 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
