

Compact zinc finger base editors that edit mitochondrial or nuclear DNA in vitro and in vivo
- PMID: 36418298
- PMCID: PMC9684478
- DOI: 10.1038/s41467-022-34784-7
Compact zinc finger base editors that edit mitochondrial or nuclear DNA in vitro and in vivo
Abstract
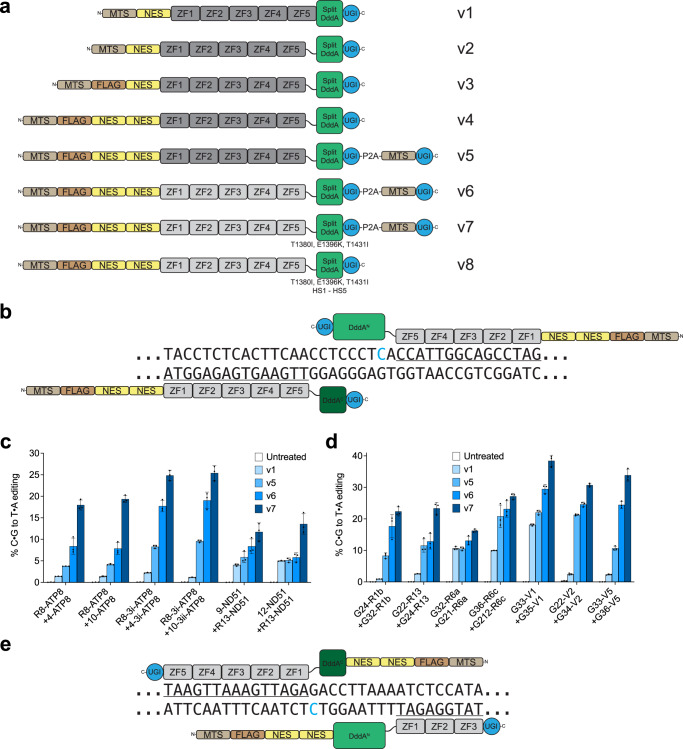
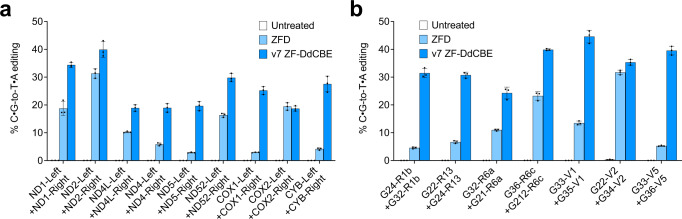
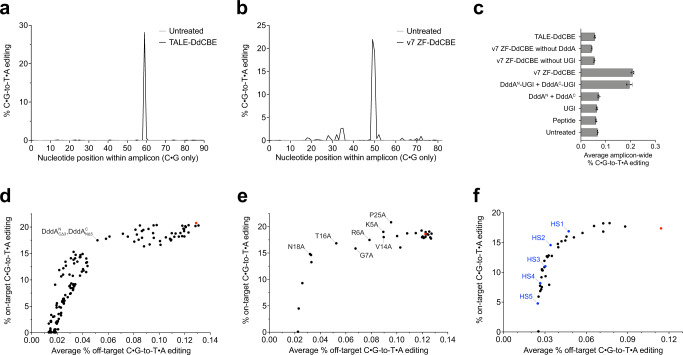
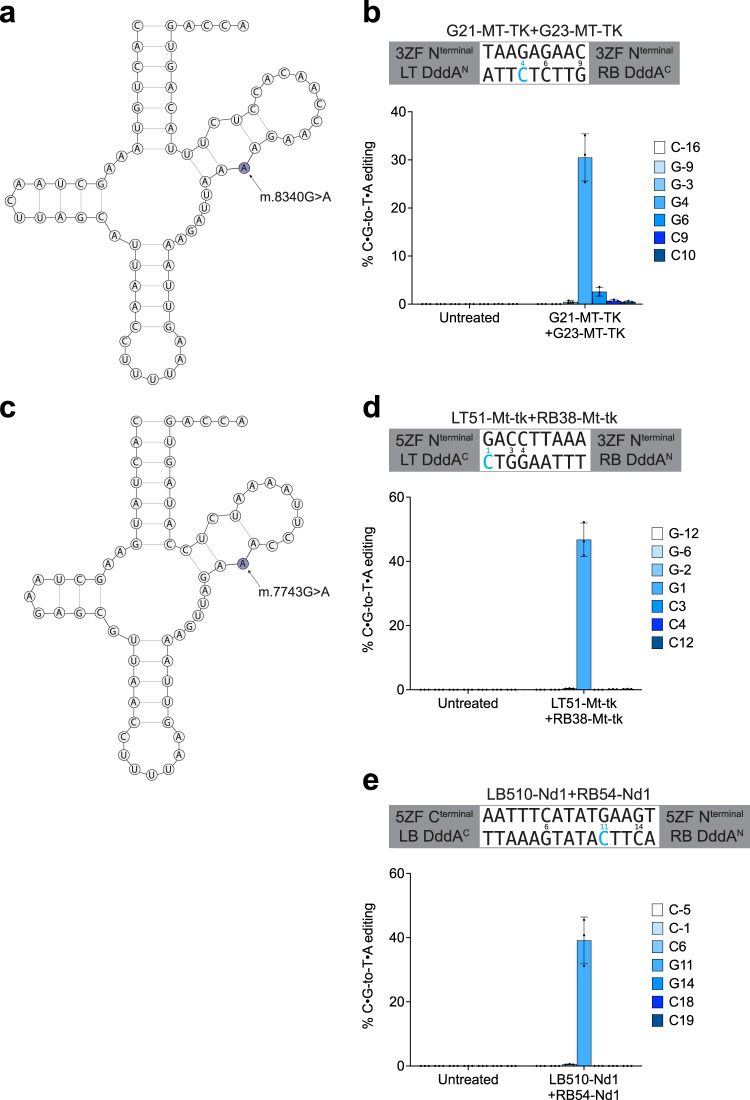
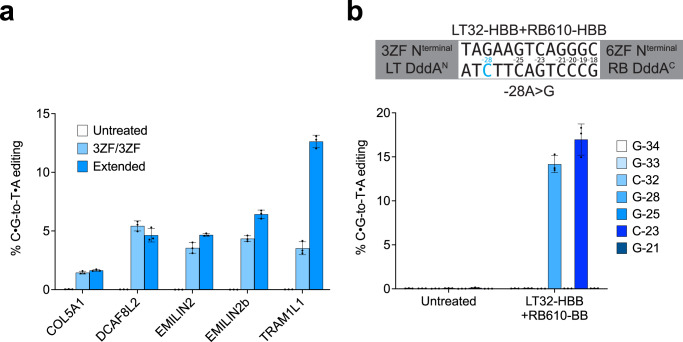
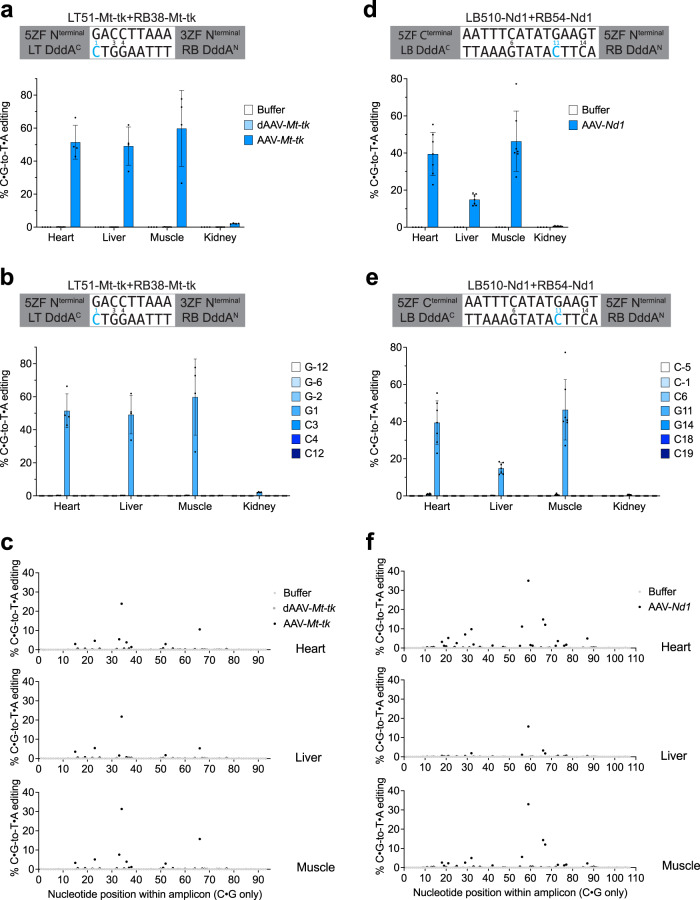
DddA-derived cytosine base editors (DdCBEs) use programmable DNA-binding TALE repeat arrays, rather than CRISPR proteins, a split double-stranded DNA cytidine deaminase (DddA), and a uracil glycosylase inhibitor to mediate C•G-to-T•A editing in nuclear and organelle DNA. Here we report the development of zinc finger DdCBEs (ZF-DdCBEs) and the improvement of their editing performance through engineering their architectures, defining improved ZF scaffolds, and installing DddA activity-enhancing mutations. We engineer variants with improved DNA specificity by integrating four strategies to reduce off-target editing. We use optimized ZF-DdCBEs to install or correct disease-associated mutations in mitochondria and in the nucleus. Leveraging their small size, we use a single AAV9 to deliver into heart, liver, and skeletal muscle in post-natal mice ZF-DdCBEs that efficiently install disease-associated mutations. While off-target editing of ZF-DdCBEs is likely too high for therapeutic applications, these findings demonstrate a compact, all-protein base editing research tool for precise editing of organelle or nuclear DNA without double-strand DNA breaks.
© 2022. The Author(s).
Conflict of interest statement
The authors declare competing financial interests: J.C.W.W. and D.R.L. have filed patent applications on this work. D.R.L. is a consultant for Prime Medicine, Beam Therapeutics, Pairwise Plants, Chroma Medicine, and Nvelop Therapeutics, companies that use or deliver genome editing or genome engineering agents, and owns equity in these companies. M.M. is listed as an inventor on a patent related to this work (WO2007071962A1) and filed a patent application on this work (PCT/GB2019/050808). M.M. is a co-founder, shareholder and Scientific Advisory Board member of Pretzel Therapeutics. P.S.-P. is a consultant for Pretzel Therapeutics. L.W. declares no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
