

Structured cerebellar connectivity supports resilient pattern separation
- PMID: 36418404
- PMCID: PMC10324966
- DOI: 10.1038/s41586-022-05471-w
Structured cerebellar connectivity supports resilient pattern separation
Erratum in
-
Publisher Correction: Structured cerebellar connectivity supports resilient pattern separation.Nature. 2023 Feb;614(7946):E18. doi: 10.1038/s41586-023-05703-7. Nature. 2023. PMID: 36631615 No abstract available.
Abstract
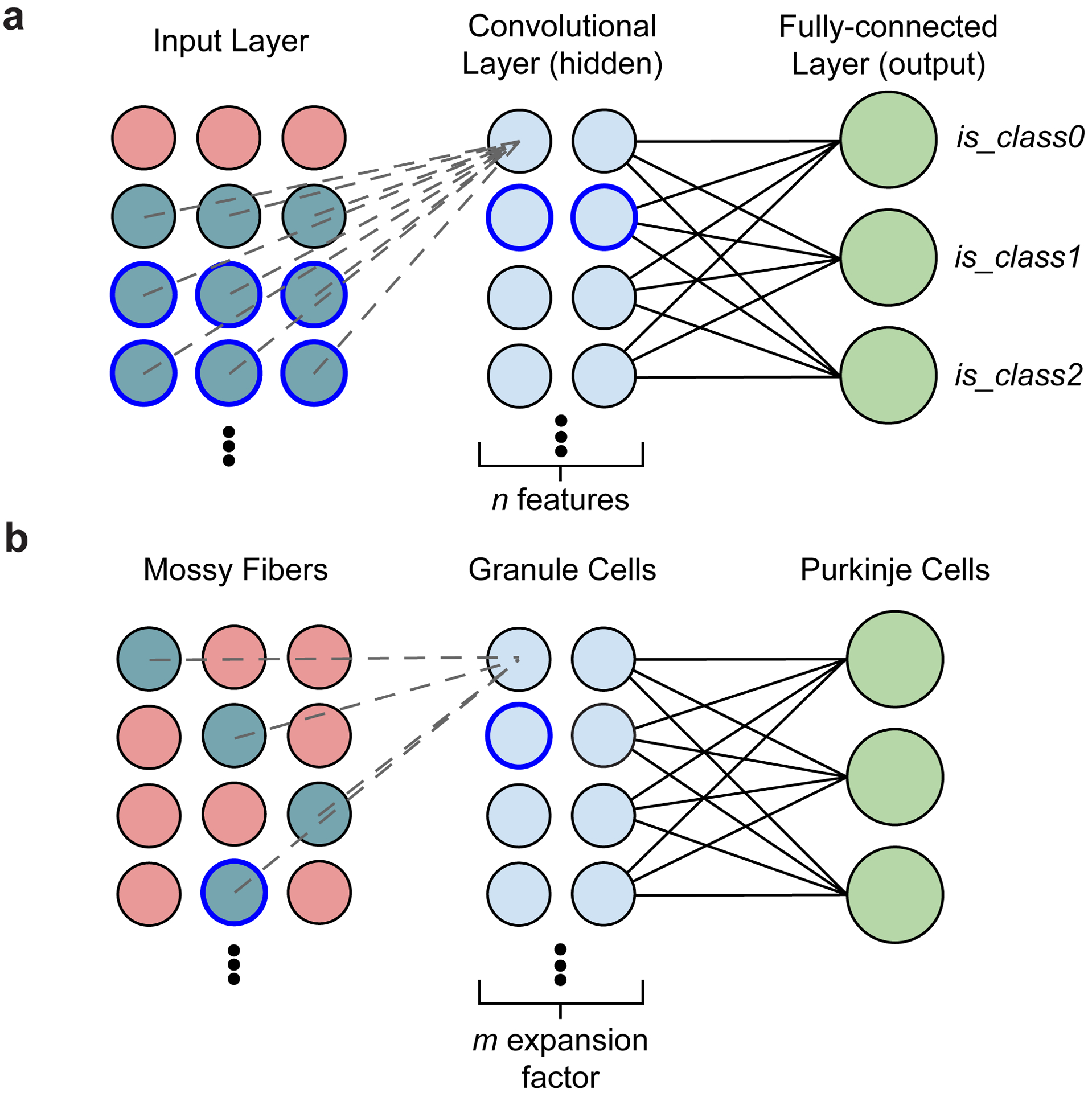
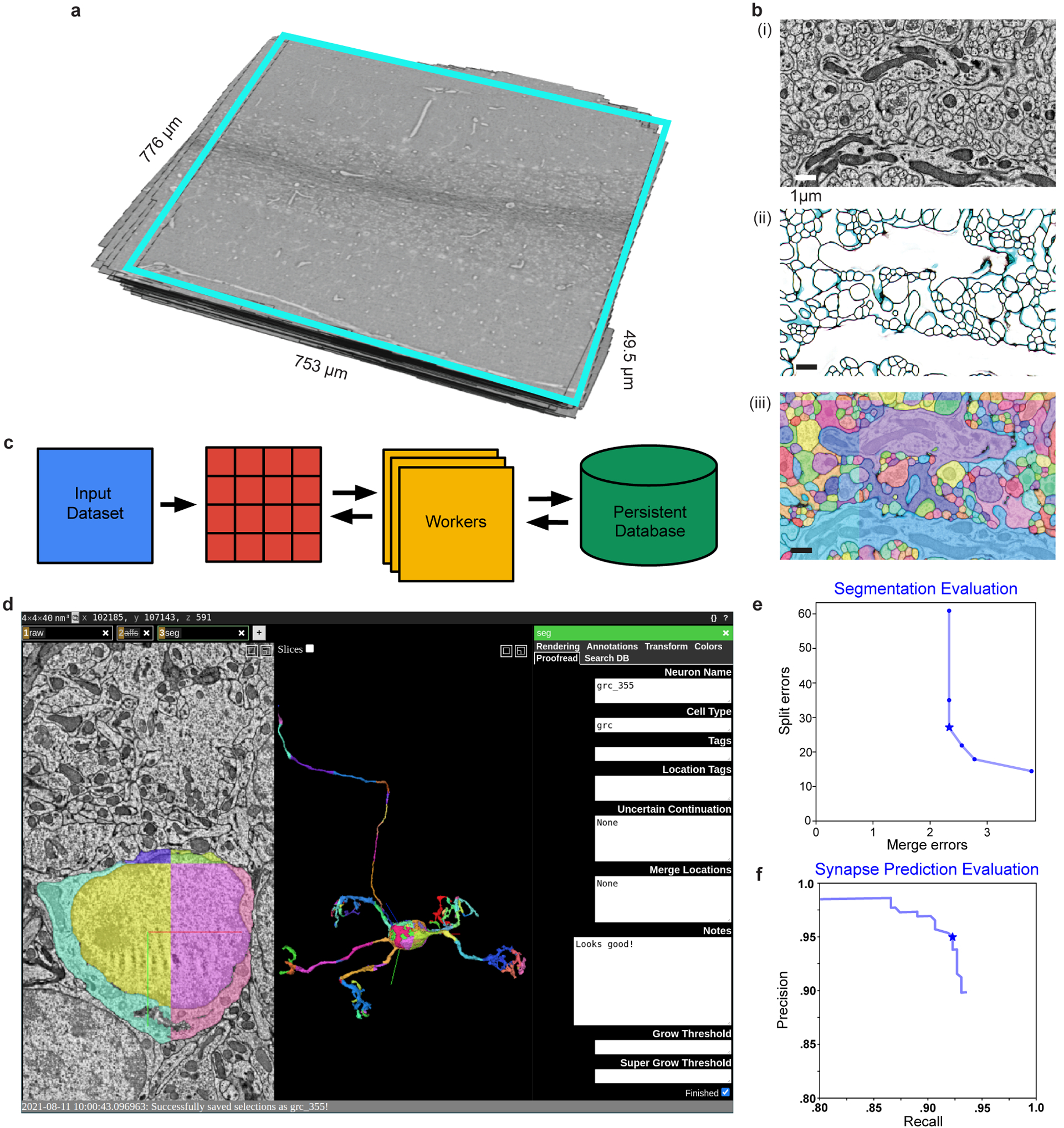
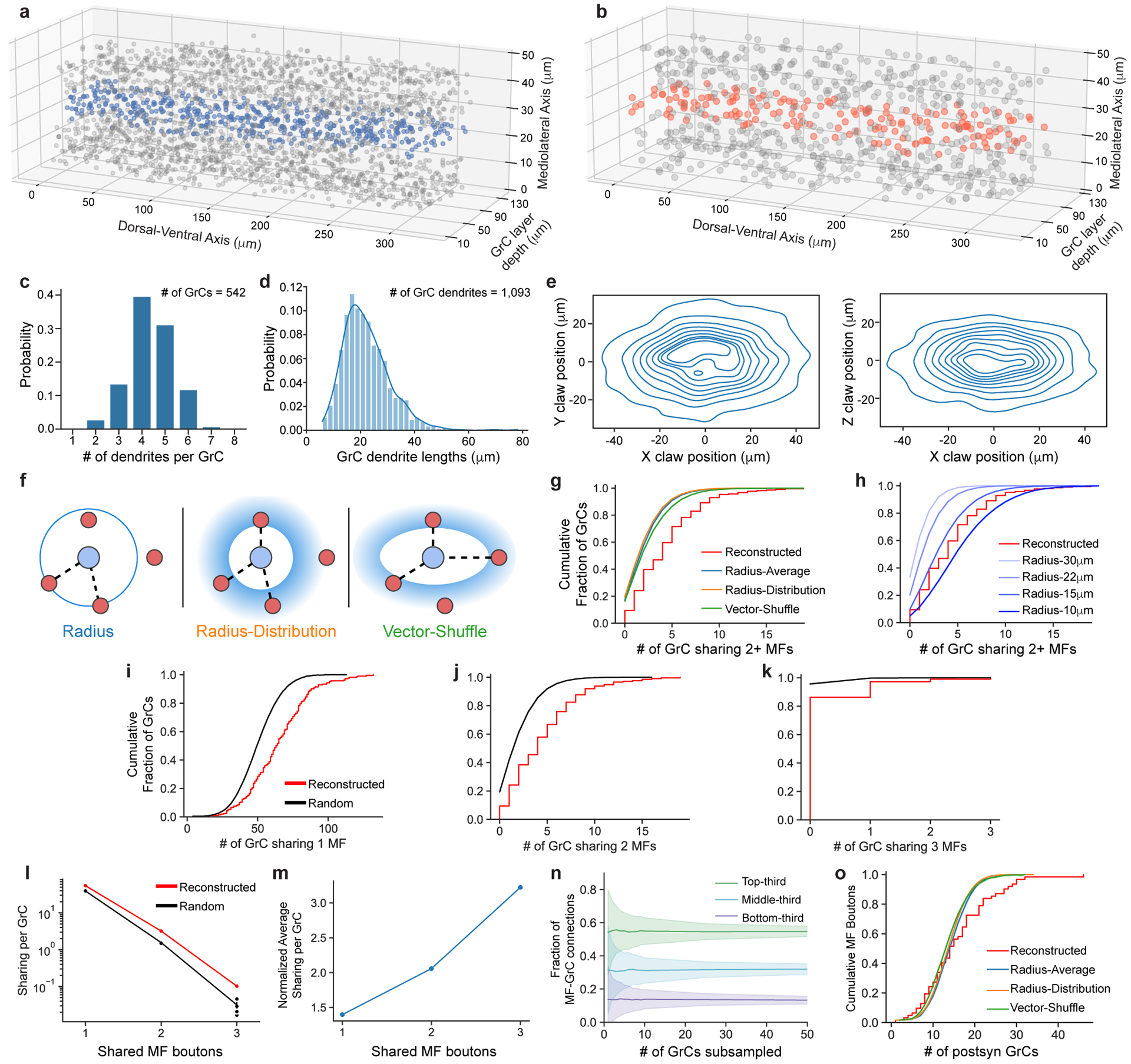
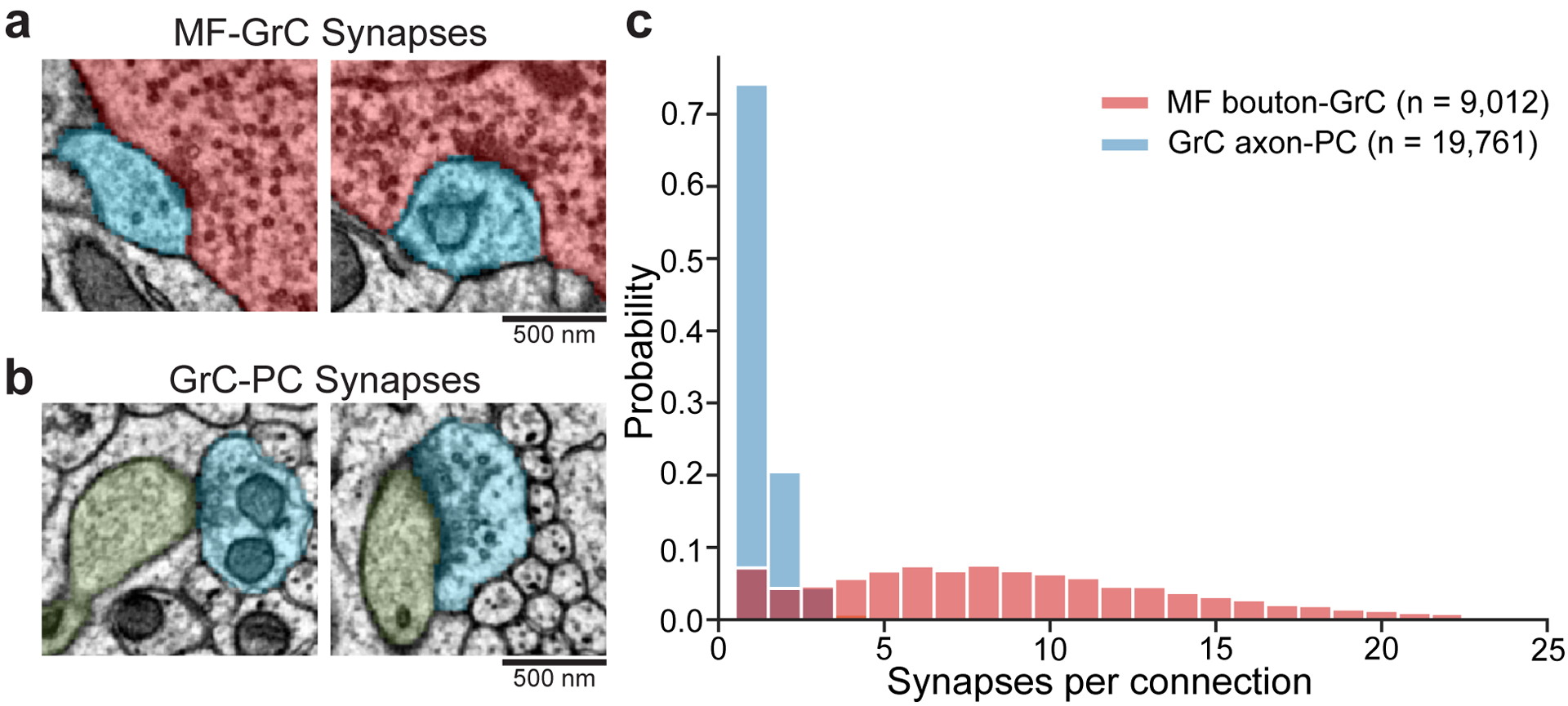
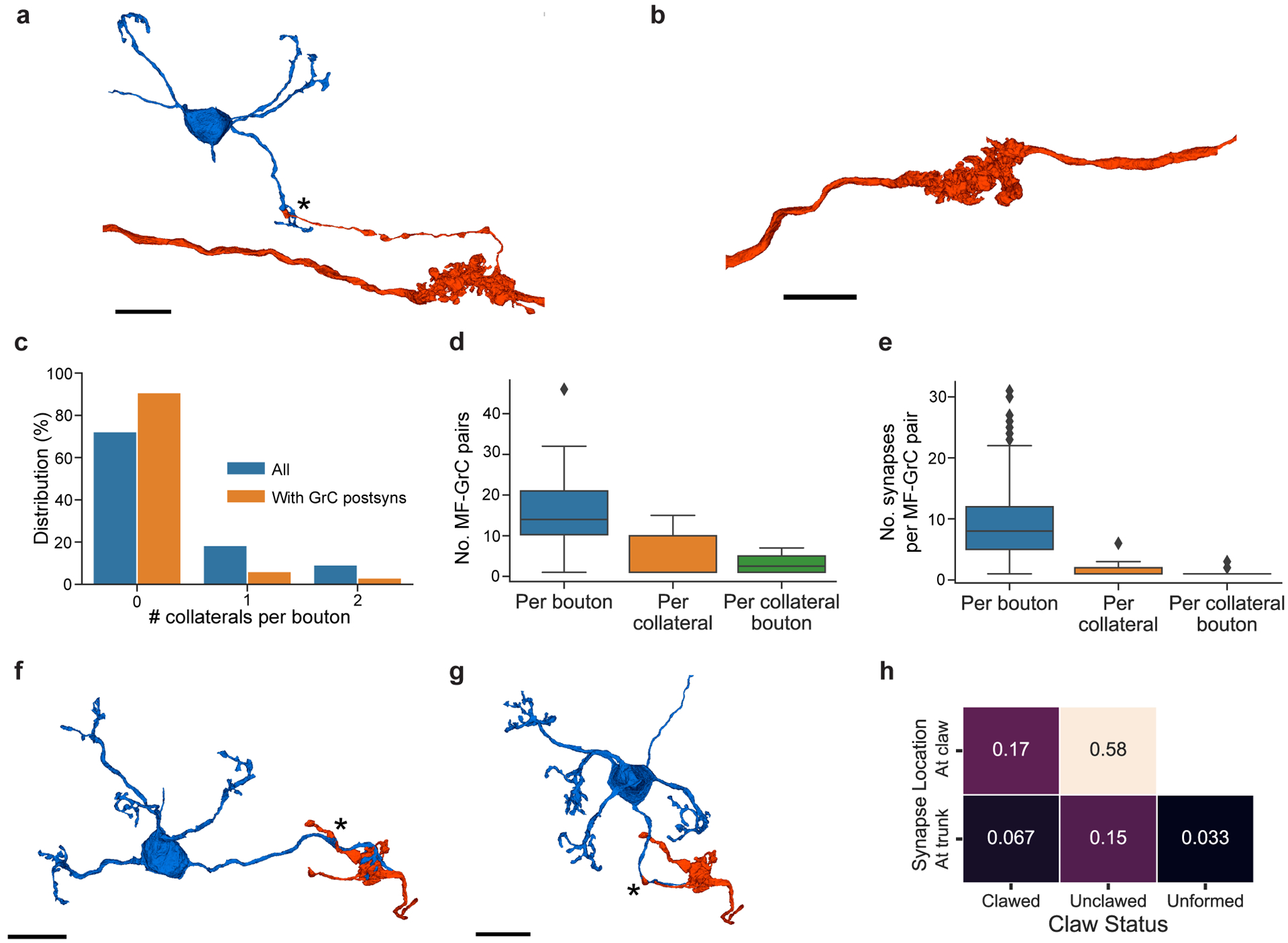
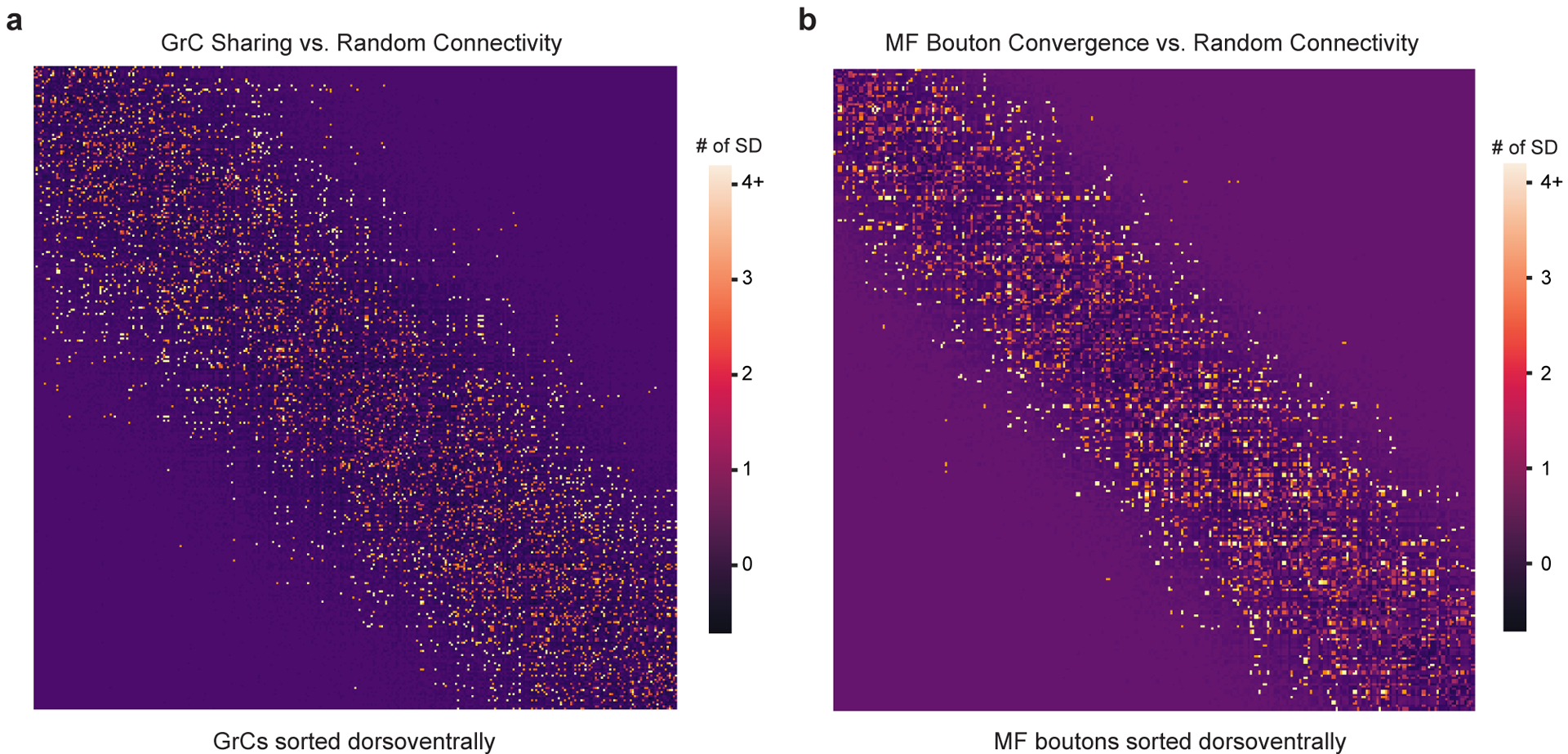
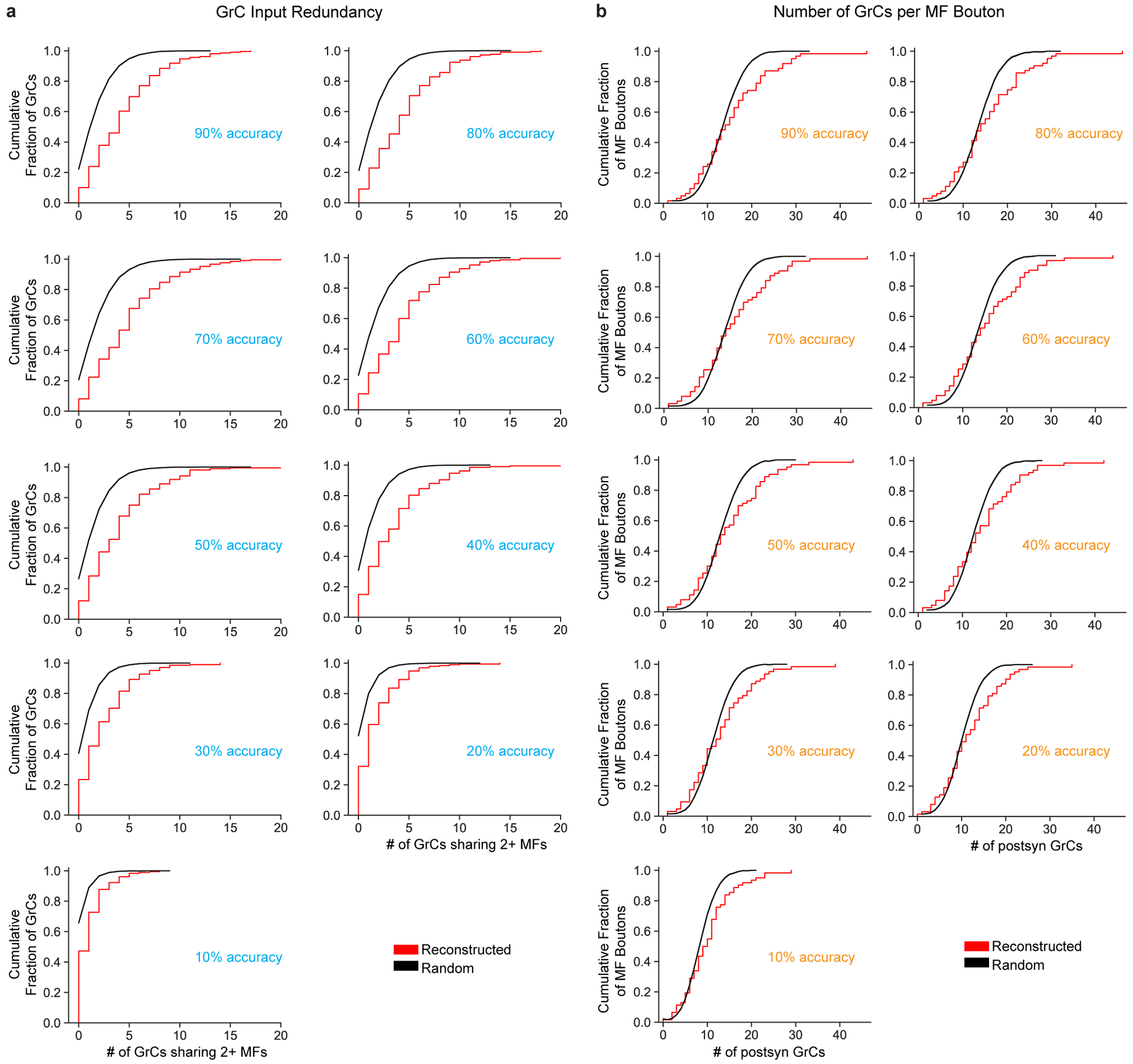
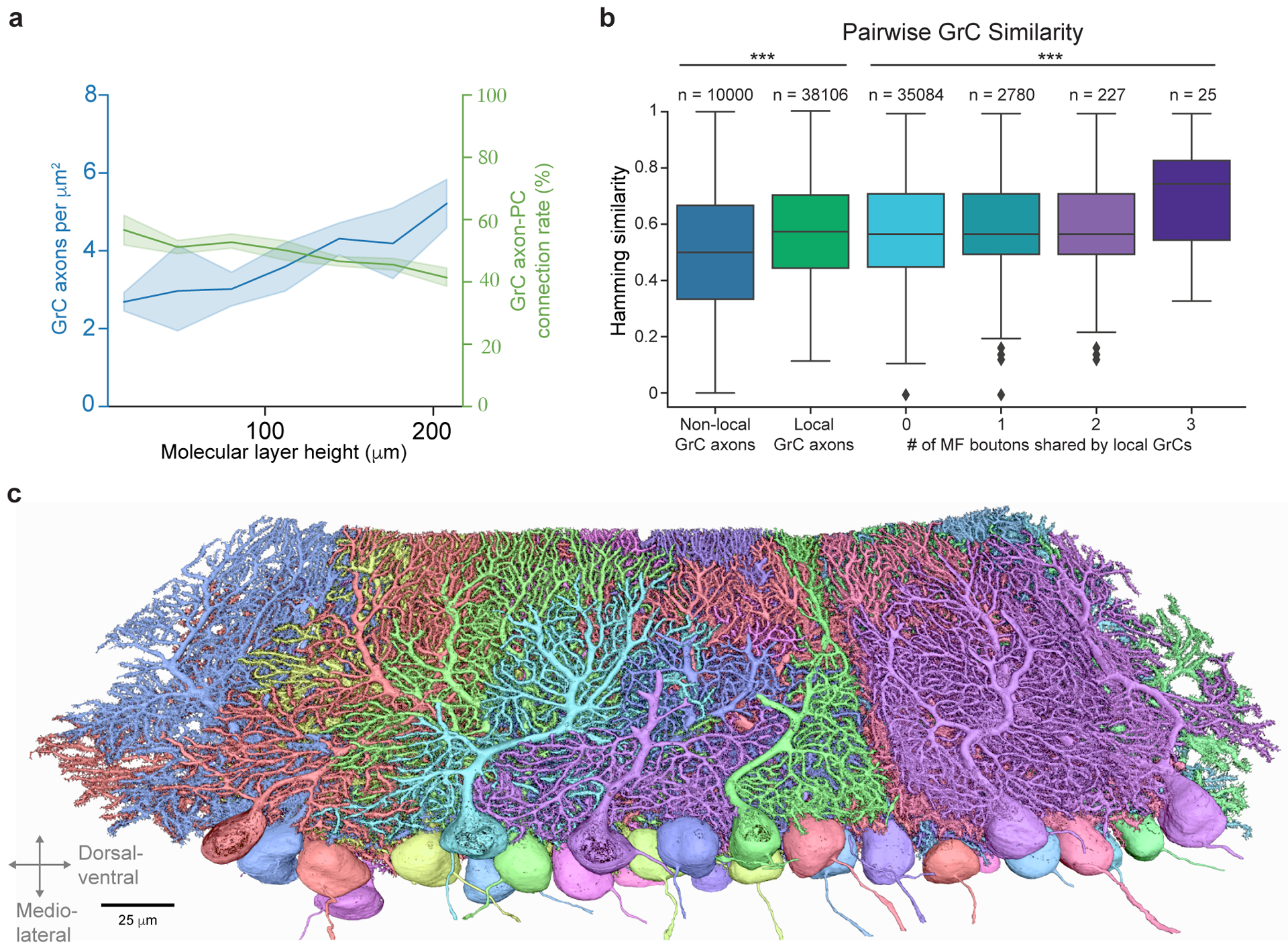
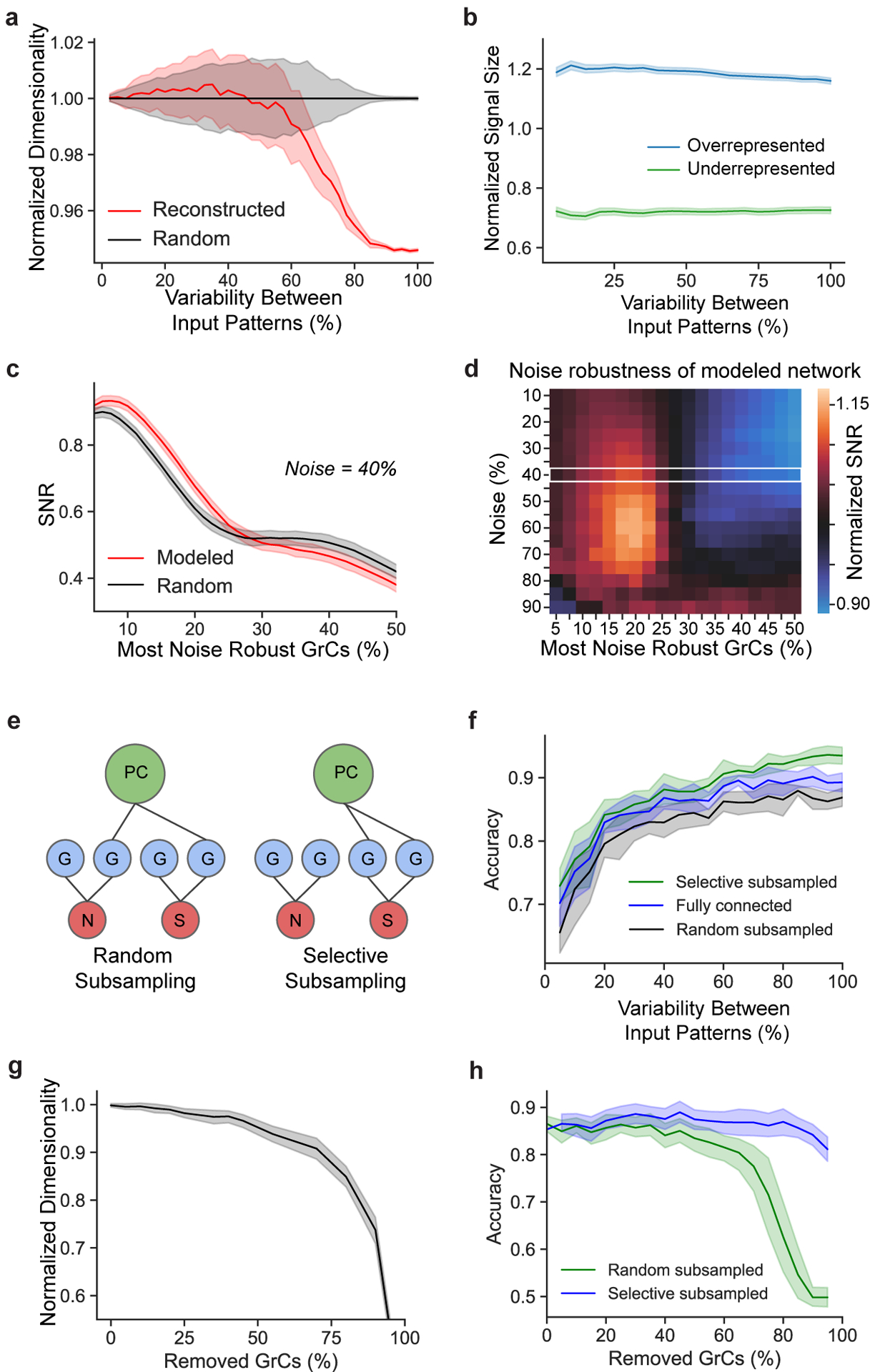
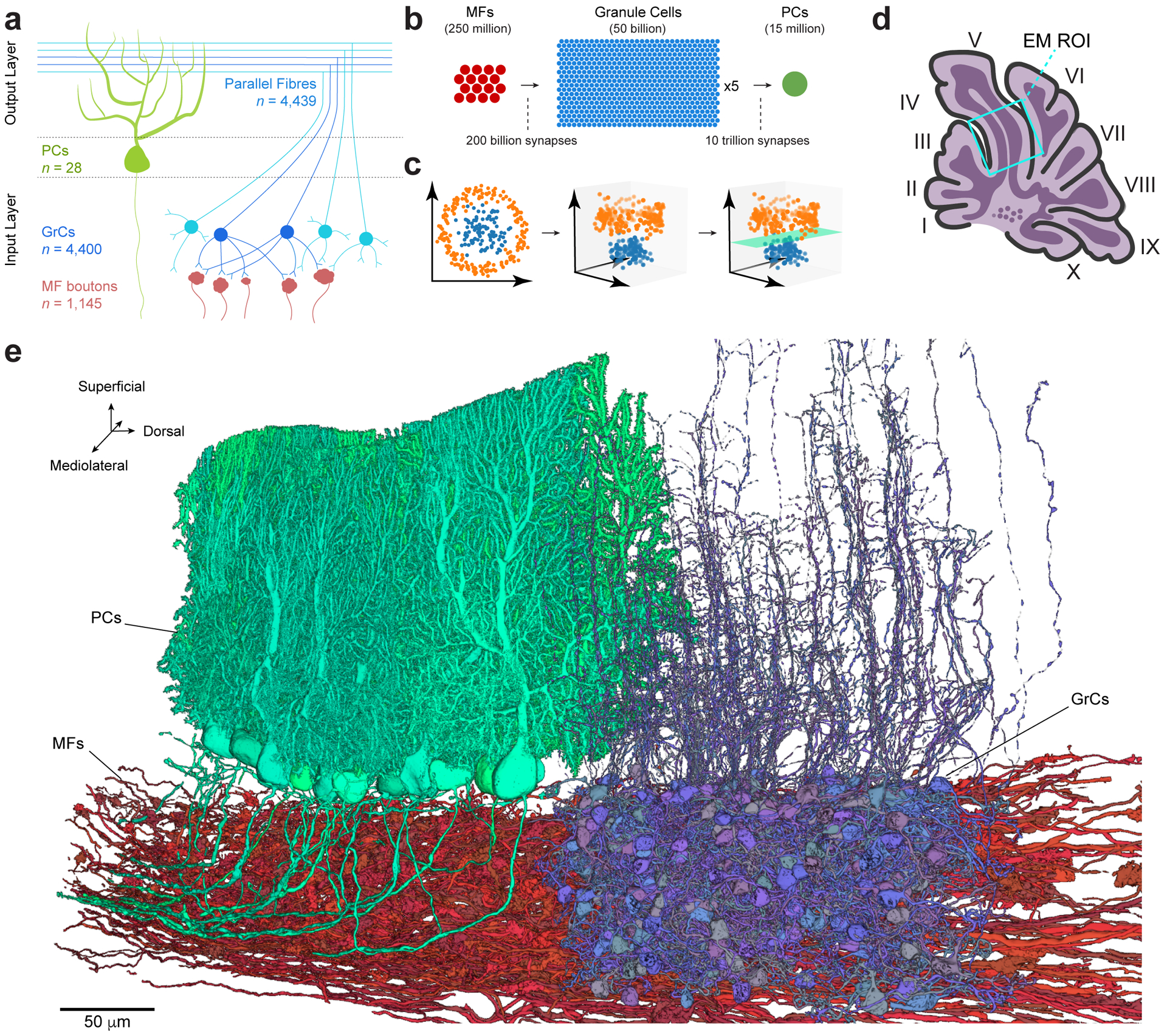
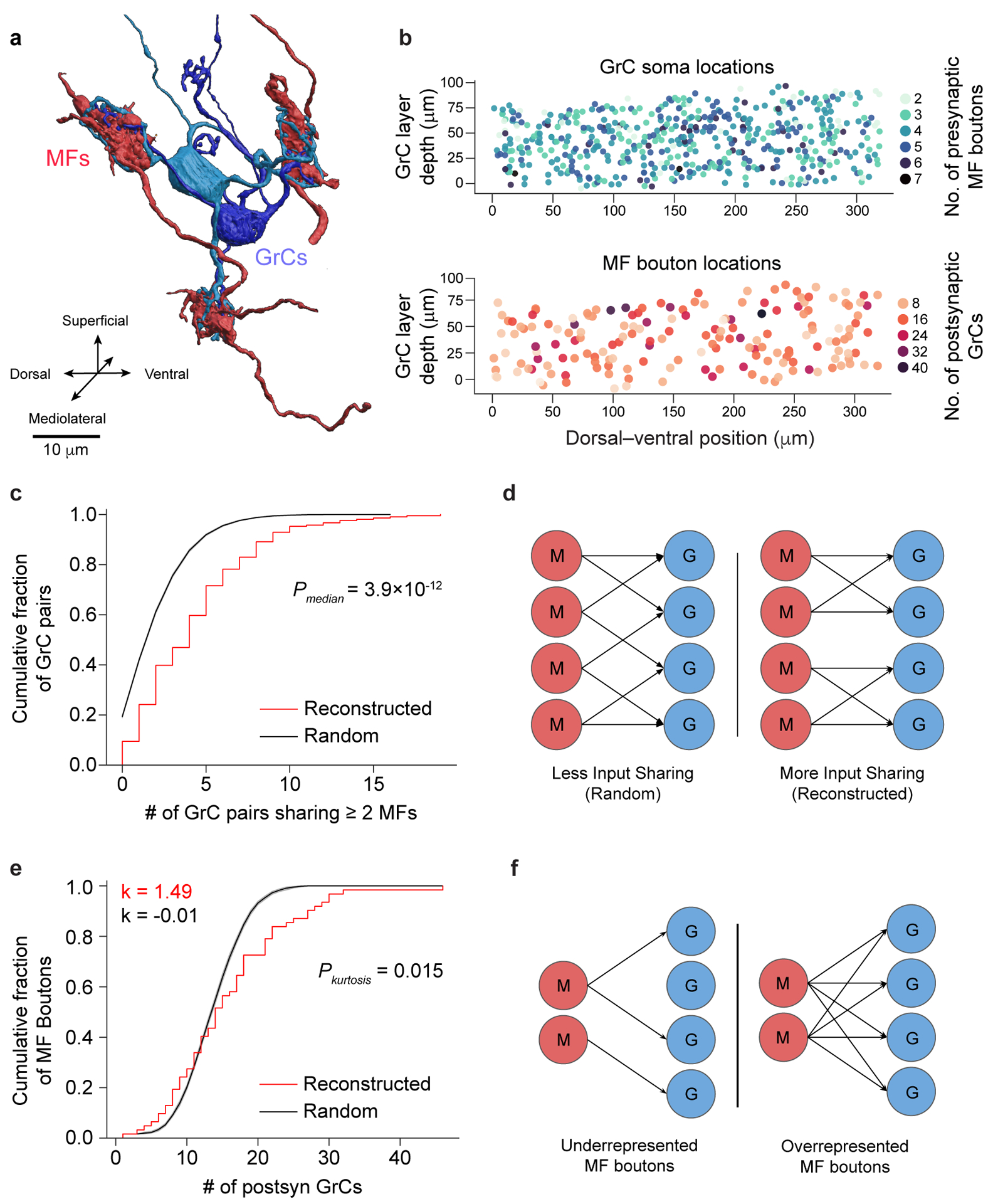
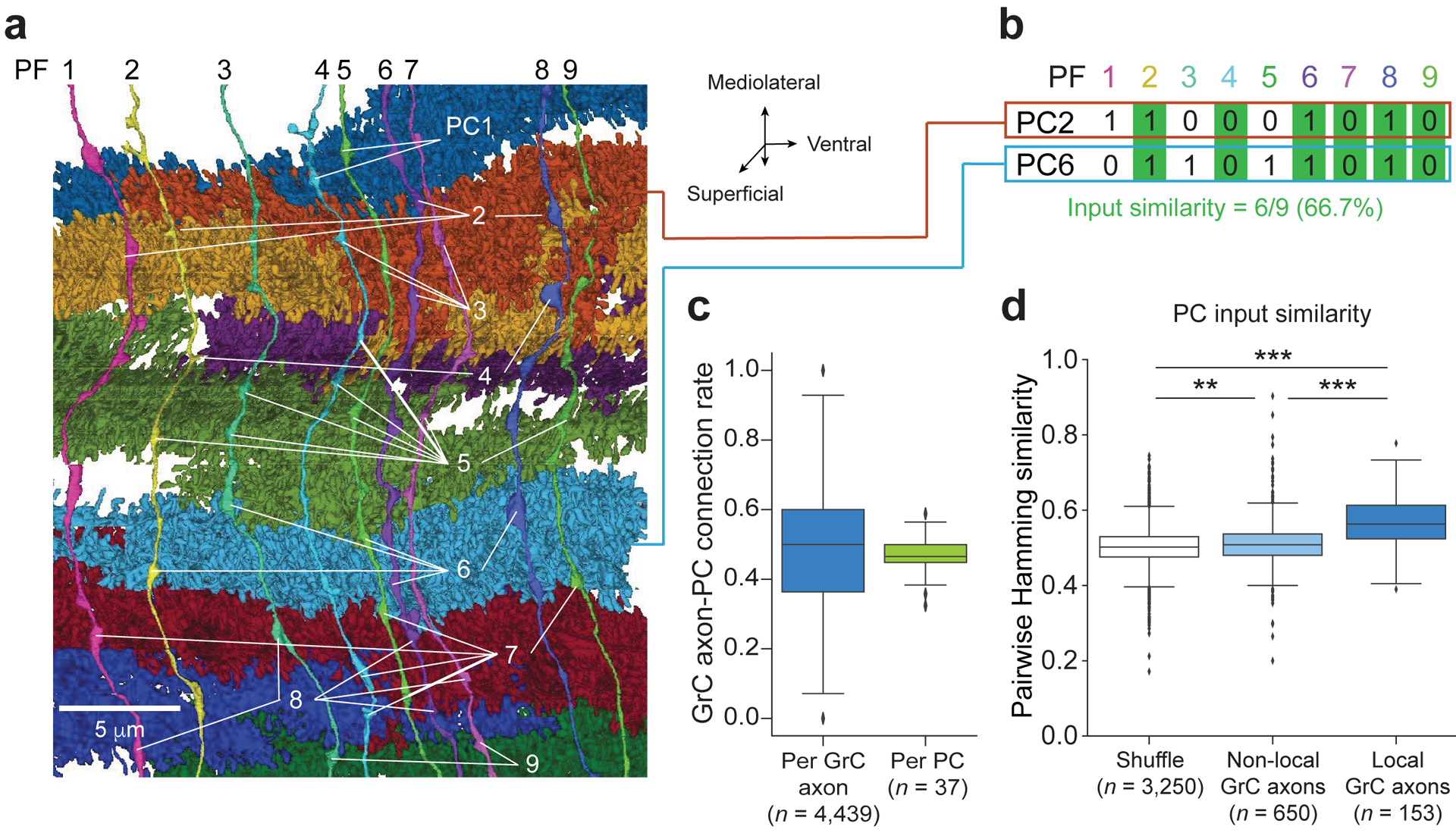
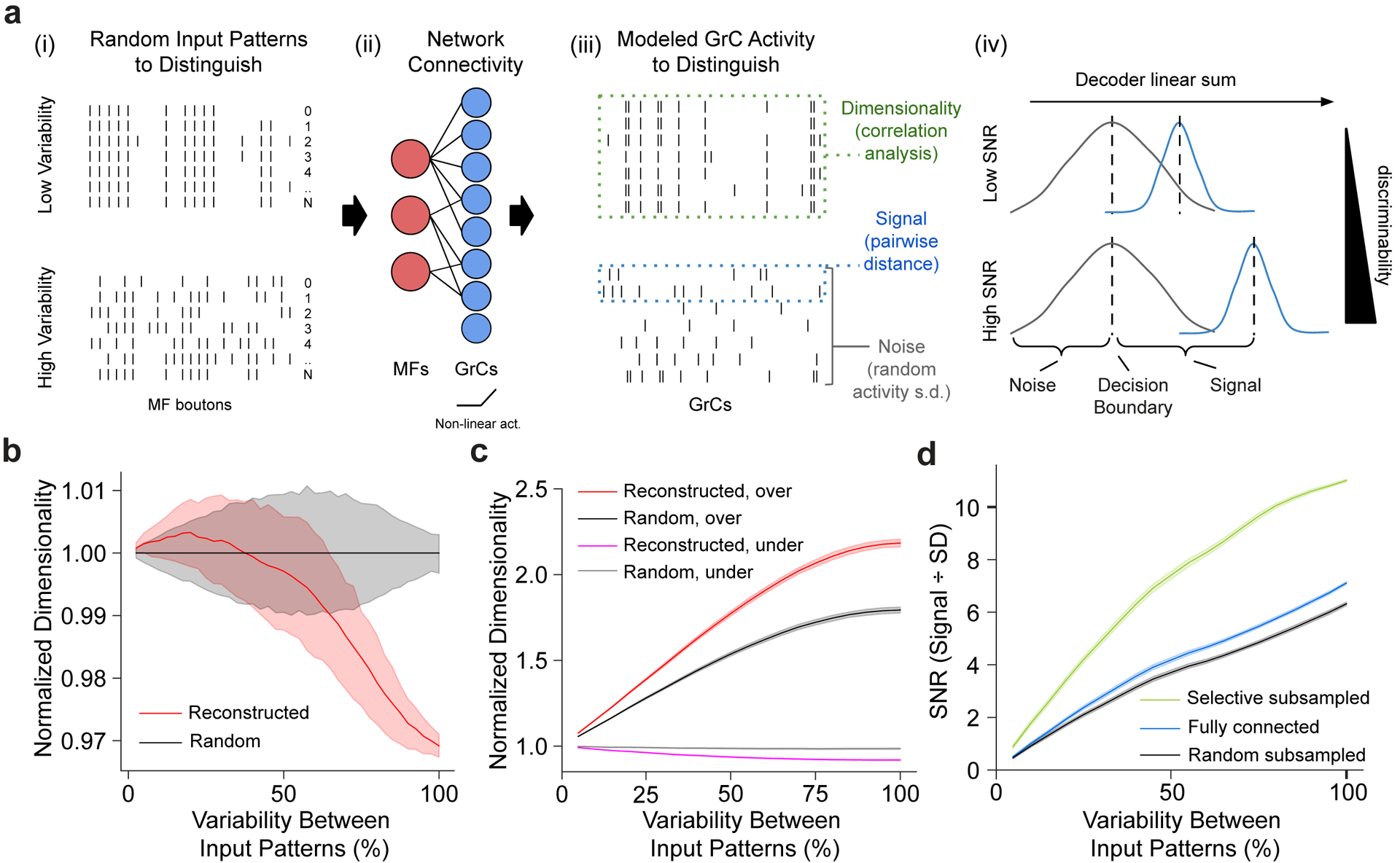
The cerebellum is thought to help detect and correct errors between intended and executed commands1,2 and is critical for social behaviours, cognition and emotion3-6. Computations for motor control must be performed quickly to correct errors in real time and should be sensitive to small differences between patterns for fine error correction while being resilient to noise7. Influential theories of cerebellar information processing have largely assumed random network connectivity, which increases the encoding capacity of the network's first layer8-13. However, maximizing encoding capacity reduces the resilience to noise7. To understand how neuronal circuits address this fundamental trade-off, we mapped the feedforward connectivity in the mouse cerebellar cortex using automated large-scale transmission electron microscopy and convolutional neural network-based image segmentation. We found that both the input and output layers of the circuit exhibit redundant and selective connectivity motifs, which contrast with prevailing models. Numerical simulations suggest that these redundant, non-random connectivity motifs increase the resilience to noise at a negligible cost to the overall encoding capacity. This work reveals how neuronal network structure can support a trade-off between encoding capacity and redundancy, unveiling principles of biological network architecture with implications for the design of artificial neural networks.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
W.C.A.L. and D.G.C.H. declare the following competing interest: Harvard University filed a patent application regarding GridTape (WO2017184621A1) on behalf of the inventors including W.C.A.L. and D.G.C.H., and negotiated licensing agreements with interested partners.
Figures
References
-
- Wolpert DM, Miall RC & Kawato M Internal models in the cerebellum. Trends Cogn. Sci 2, 338–347 (1998). - PubMed
-
- Strick PL, Dum RP & Fiez JA Cerebellum and nonmotor function. Annu. Rev. Neurosci 32, 413–434 (2009). - PubMed
-
- Schmahmann JD Disorders of the cerebellum: ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J. Neuropsychiatry Clin. Neurosci 16, 367–378 (2004). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
