

Spliceosomal profiling identifies EIF4A3 as a novel oncogene in hepatocellular carcinoma acting through the modulation of FGFR4 splicing
- PMID: 36419260
- PMCID: PMC9684617
- DOI: 10.1002/ctm2.1102
Spliceosomal profiling identifies EIF4A3 as a novel oncogene in hepatocellular carcinoma acting through the modulation of FGFR4 splicing
Abstract
Introduction: Altered splicing landscape is an emerging cancer hallmark; however, the dysregulation and implication of the cellular machinery controlling this process (spliceosome components and splicing factors) in hepatocellular carcinoma (HCC) is poorly known. This study aimed to comprehensively characterize the spliceosomal profile and explore its role in HCC.
Methods: Expression levels of 70 selected spliceosome components and splicing factors and clinical implications were evaluated in two retrospective and six in silico HCC cohorts. Functional, molecular and mechanistic studies were implemented in three cell lines (HepG2, Hep3B and SNU-387) and preclinical Hep3B-induced xenograft tumours.
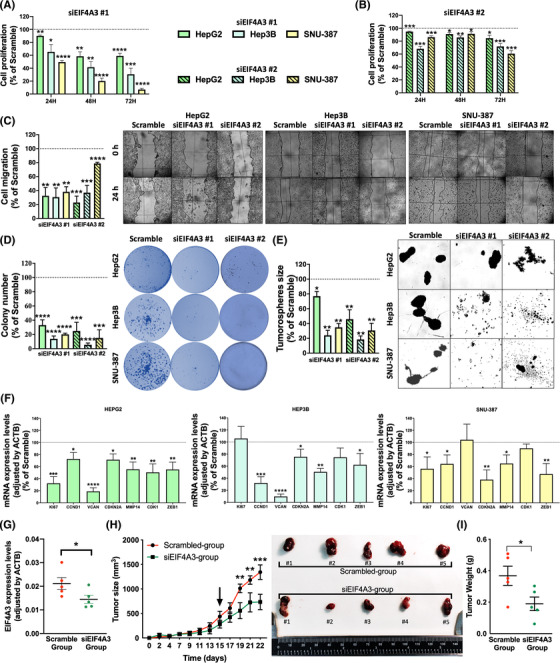
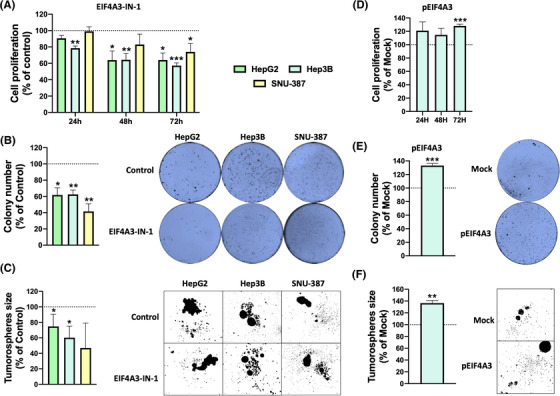
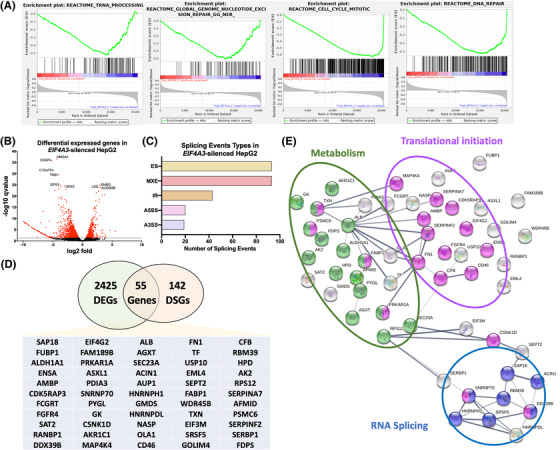
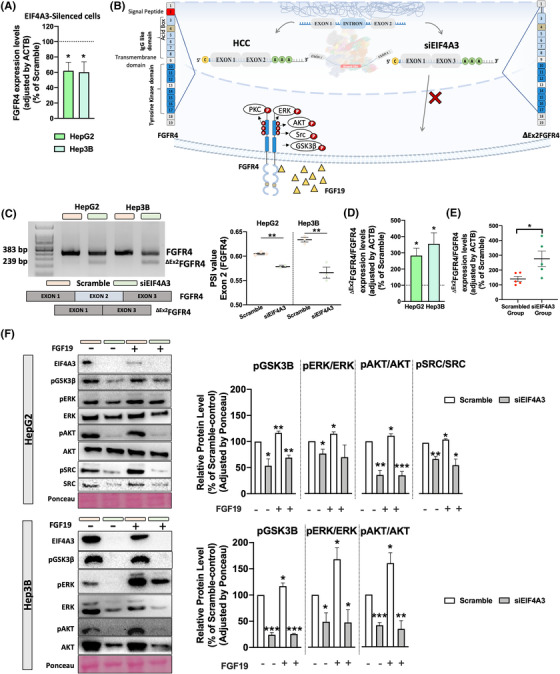
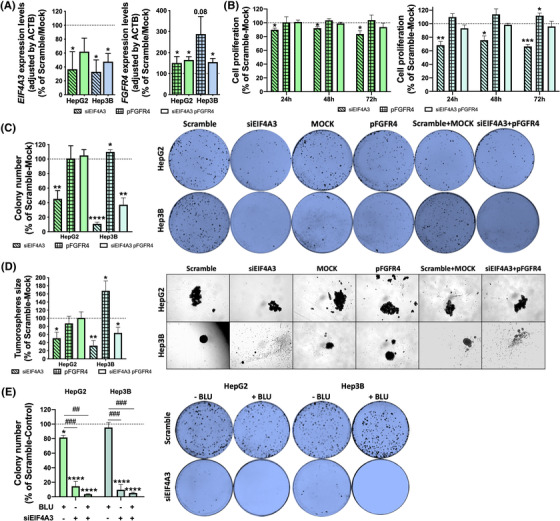
Results: Spliceosomal dysregulations were consistently found in retrospective and in silico cohorts. EIF4A3, RBM3, ESRP2 and SRPK1 were the most dysregulated spliceosome elements in HCC. EIF4A3 expression was associated with decreased survival and greater recurrence. Plasma EIF4A3 levels were significantly elevated in HCC patients. In vitro EIF4A3-silencing (or pharmacological inhibition) resulted in reduced aggressiveness, and hindered xenograft-tumours growth in vivo, whereas EIF4A3 overexpression increased tumour aggressiveness. EIF4A3-silencing altered the expression and splicing of key HCC-related genes, specially FGFR4. EIF4A3-silencing blocked the cellular response to the natural ligand of FGFR4, FGF19. Functional consequences of EIF4A3-silencing were mediated by FGFR4 splicing as the restoration of non-spliced FGFR4 full-length version blunted these effects, and FGFR4 inhibition did not exert further effects in EIF4A3-silenced cells.
Conclusions: Splicing machinery is strongly dysregulated in HCC, providing a source of new diagnostic, prognostic and therapeutic options in HCC. EIF4A3 is consistently elevated in HCC patients and associated with tumour aggressiveness and mortality, through the modulation of FGFR4 splicing.
Keywords: FGF19; liver cancer; preclinical model; splicing machinery.
© 2022 The Authors. Clinical and Translational Medicine published by John Wiley & Sons Australia, Ltd on behalf of Shanghai Institute of Clinical Bioinformatics.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures


References
-
- Villanueva A. Hepatocellular carcinoma. N Engl J Med. 2019;380(15):1450‐1462. - PubMed
-
- Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71:209‐249. - PubMed
-
- Wang E, Aifantis I. RNA splicing and cancer. Trends Cancer. 2020;6(8):631‐644. - PubMed
-
- Castillo J, Goni S, Latasa MU, et al. Amphiregulin induces the alternative splicing of p73 into its oncogenic isoform DeltaEx2p73 in human hepatocellular tumors. Gastroenterology. 2009;137(5):1805‐1815.e1‐4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous