

ABA signaling prevents phosphodegradation of the SR45 splicing factor to alleviate inhibition of early seedling development in Arabidopsis
- PMID: 36419364
- PMCID: PMC10030365
- DOI: 10.1016/j.xplc.2022.100495
ABA signaling prevents phosphodegradation of the SR45 splicing factor to alleviate inhibition of early seedling development in Arabidopsis
Abstract
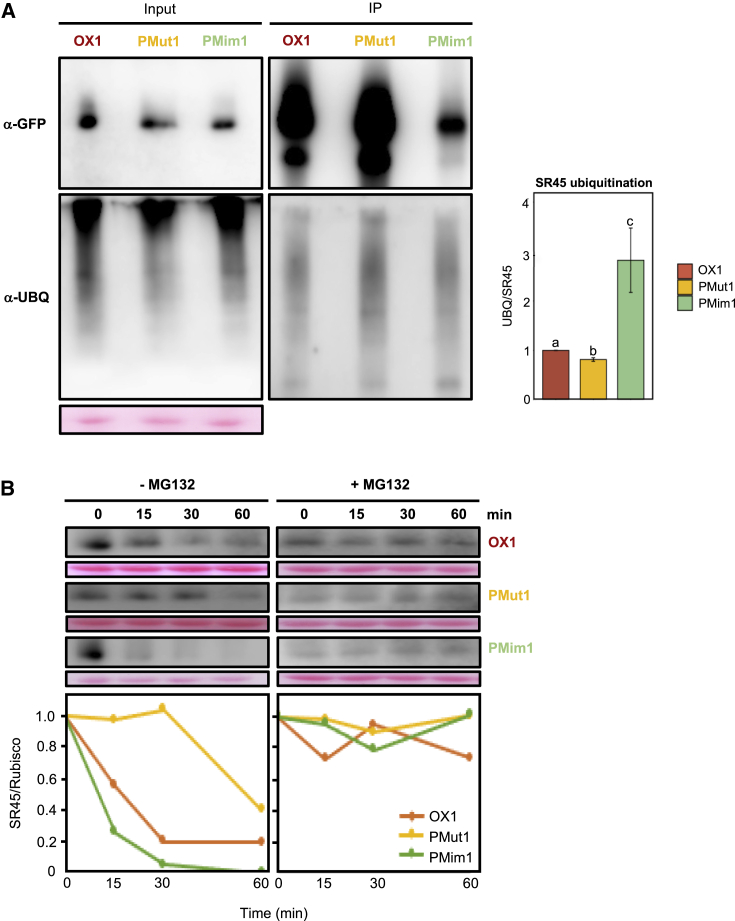
Serine/arginine-rich (SR) proteins are conserved splicing regulators that play important roles in plant stress responses, namely those mediated by the abscisic acid (ABA) hormone. The Arabidopsis thaliana SR-like protein SR45 is a described negative regulator of the ABA pathway during early seedling development. How the inhibition of growth by ABA signaling is counteracted to maintain plant development under stress conditions remains largely unknown. Here, we show that SR45 overexpression reduces Arabidopsis sensitivity to ABA during early seedling development. Biochemical and confocal microscopy analyses of transgenic plants expressing fluorescently tagged SR45 revealed that exposure to ABA dephosphorylates the protein at multiple amino acid residues and leads to its accumulation, due to SR45 stabilization via reduced ubiquitination and proteasomal degradation. Using phosphomutant and phosphomimetic transgenic Arabidopsis lines, we demonstrate the functional relevance of ABA-mediated dephosphorylation of a single SR45 residue, T264, in antagonizing SR45 ubiquitination and degradation to promote its function as a repressor of seedling ABA sensitivity. Our results reveal a mechanism that negatively autoregulates ABA signaling and allows early plant growth under stress via posttranslational control of the SR45 splicing factor.
Keywords: Arabidopsis thaliana; SR proteins; abscisic acid; alternative splicing; protein phosphorylation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Figures






References
-
- Al-Sady B., Ni W., Kircher S., Schäfer E., Quail P.H. Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol. Cell. 2006;23:439–446. - PubMed
-
- Albaqami M., Laluk K., Reddy A.S.N. The Arabidopsis splicing regulator SR45 confers salt tolerance in a splice isoform-dependent manner. Plant Mol. Biol. 2019;100:379–390. - PubMed
-
- Bhaskara G.B., Wong M.M., Verslues P.E. The flip side of phospho-signalling: regulation of protein dephosphorylation and the protein phosphatase 2Cs. Plant Cell Environ. 2019;42:2913–2930. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
