

Molecular determinants of the interaction between HSV-1 glycoprotein D and heparan sulfate
- PMID: 36419932
- PMCID: PMC9678342
- DOI: 10.3389/fmolb.2022.1043713
Molecular determinants of the interaction between HSV-1 glycoprotein D and heparan sulfate
Abstract
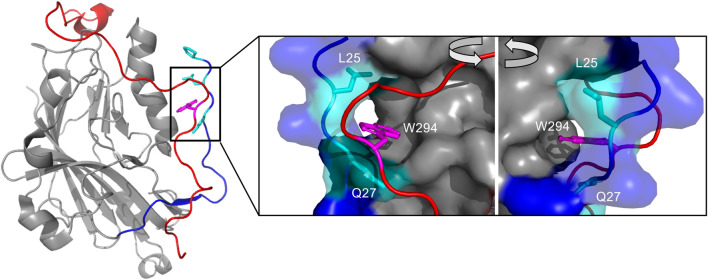
Literature has well-established the importance of 3-O-sulfation of neuronal cell surface glycan heparan sulfate (HS) to its interaction with herpes simplex virus type 1 glycoprotein D (gD). Previous investigations of gD to its viral receptors HVEM and nectin-1 also highlighted the conformational dynamics of gD's N- and C-termini, necessary for viral membrane fusion. However, little is known on the structural interactions of gD with HS. Here, we present our findings on this interface from both the glycan and the protein perspective. We used C-terminal and N-terminal gD variants to probe the role of their respective regions in gD/HS binding. The N-terminal truncation mutants (with Δ1-22) demonstrate equivalent or stronger binding to heparin than their intact glycoproteins, indicating that the first 22 amino acids are disposable for heparin binding. Characterization of the conformational differences between C-terminal truncated mutants by sedimentation velocity analytical ultracentrifugation distinguished between the "open" and "closed" conformations of the glycoprotein D, highlighting the region's modulation of receptor binding. From the glycan perspective, we investigated gD interacting with heparin, heparan sulfate, and other de-sulfated and chemically defined oligosaccharides using surface plasmon resonance and glycan microarray. The results show a strong preference of gD for 6-O-sulfate, with 2-O-sulfation becoming more important in the presence of 6-O-S. Additionally, 3-O-sulfation shifted the chain length preference of gD from longer chain to mid-chain length, reaffirming the sulfation site's importance to the gD/HS interface. Our results shed new light on the molecular details of one of seven known protein-glycan interactions with 3-O-sulfated heparan sulfate.
Keywords: HSV-1; glycoprotein D; heparan sulfate; heparin; herpes.
Copyright © 2022 Gandy, Canning, Lou, Xia, He, Su, Cairns, Liu, Zhang, Linhardt, Cohen and Wang.
Conflict of interest statement
JL is the founder of Glycan Therapeutics and GS is an employee of Glycan Therapeutics. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




Similar articles
-
Mutations in the N termini of herpes simplex virus type 1 and 2 gDs alter functional interactions with the entry/fusion receptors HVEM, nectin-2, and 3-O-sulfated heparan sulfate but not with nectin-1.J Virol. 2003 Sep;77(17):9221-31. doi: 10.1128/jvi.77.17.9221-9231.2003. J Virol. 2003. PMID: 12915538 Free PMC article.
-
Participation of 3-O-sulfated heparan sulfates in the protection of macrophages by herpes simplex virus-1 glycoprotein D and cyclophilin B against apoptosis.FEBS Open Bio. 2016 Dec 24;7(2):133-148. doi: 10.1002/2211-5463.12145. eCollection 2017 Feb. FEBS Open Bio. 2016. PMID: 28174681 Free PMC article.
-
Kinetic analysis of glycoprotein C of herpes simplex virus types 1 and 2 binding to heparin, heparan sulfate, and complement component C3b.Virology. 2002 Mar 15;294(2):324-32. doi: 10.1006/viro.2001.1326. Virology. 2002. PMID: 12009874
-
Archaic connectivity between the sulfated heparan sulfate and the herpesviruses - An evolutionary potential for cross-species interactions.Comput Struct Biotechnol J. 2023 Jan 13;21:1030-1040. doi: 10.1016/j.csbj.2023.01.005. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 36733705 Free PMC article. Review.
-
Receptors and ligands for herpes simplex viruses: Novel insights for drug targeting.Drug Discov Today. 2022 Jan;27(1):185-195. doi: 10.1016/j.drudis.2021.10.004. Epub 2021 Oct 19. Drug Discov Today. 2022. PMID: 34678489 Review.
Cited by
-
Targeting heparan sulfate-protein interactions with oligosaccharides and monoclonal antibodies.Front Mol Biosci. 2023 May 19;10:1194293. doi: 10.3389/fmolb.2023.1194293. eCollection 2023. Front Mol Biosci. 2023. PMID: 37275960 Free PMC article. Review.
-
Oncolytic virotherapy against lung cancer: key receptors and signaling pathways of viral entry.Front Immunol. 2024 Oct 4;15:1473288. doi: 10.3389/fimmu.2024.1473288. eCollection 2024. Front Immunol. 2024. PMID: 39430750 Free PMC article. Review.
-
Alteration of Neuropilin-1 and Heparan Sulfate Interaction Impairs Murine B16 Tumor Growth.ACS Chem Biol. 2024 Aug 16;19(8):1820-1835. doi: 10.1021/acschembio.4c00389. Epub 2024 Aug 4. ACS Chem Biol. 2024. PMID: 39099090 Free PMC article.
-
Feline herpesvirus 1 (FHV-1) enters the cell by receptor-mediated endocytosis.J Virol. 2023 Aug 31;97(8):e0068123. doi: 10.1128/jvi.00681-23. Epub 2023 Jul 26. J Virol. 2023. PMID: 37493545 Free PMC article.
-
Mechanisms and Research Methods of Protein Modification in Virus Entry.Appl Biochem Biotechnol. 2025 Jul 19. doi: 10.1007/s12010-025-05333-x. Online ahead of print. Appl Biochem Biotechnol. 2025. PMID: 40682621 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous