

Regulation of Na+/H+ exchangers, Na+/K+ transporters, and lignin biosynthesis genes, along with lignin accumulation, sodium extrusion, and antioxidant defense, confers salt tolerance in alfalfa
- PMID: 36420040
- PMCID: PMC9676661
- DOI: 10.3389/fpls.2022.1041764
Regulation of Na+/H+ exchangers, Na+/K+ transporters, and lignin biosynthesis genes, along with lignin accumulation, sodium extrusion, and antioxidant defense, confers salt tolerance in alfalfa
Abstract
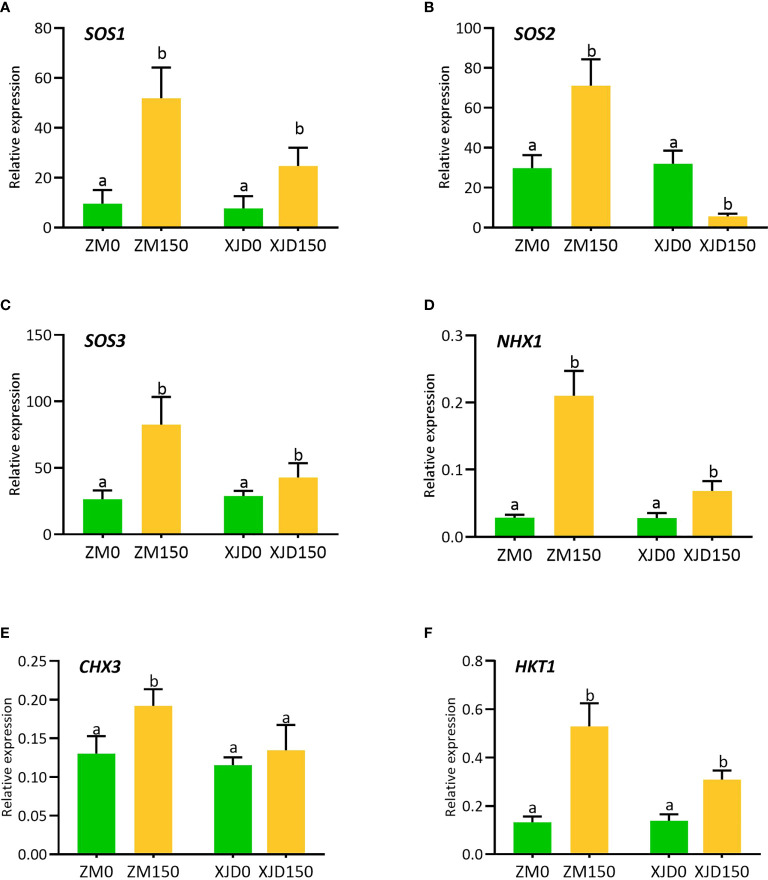
Accumulation of high sodium (Na+) leads to disruption of metabolic processes and decline in plant growth and productivity. Therefore, this study was undertaken to clarify how Na+/H+ exchangers and Na+/K+ transporter genes contribute to Na+ homeostasis and the substantial involvement of lignin biosynthesis genes in salt tolerance in alfalfa (Medicago sativa L.), which is poorly understood. In this study, high Na+ exhibited a substantial reduction of morphophysiological indices and induced oxidative stress indicators in Xingjiang Daye (XJD; sensitive genotype), while Zhongmu (ZM; tolerant genotype) remained unaffected. The higher accumulation of Na+ and the lower accumulation of K+ and K+/(Na+ + K+) ratio were found in roots and shoots of XJD compared with ZM under salt stress. The ZM genotype showed a high expression of SOS1 (salt overly sensitive 1), NHX1 (sodium/hydrogen exchanger 1), and HKT1 (high-affinity potassium transporter 1), which were involved in K+ accumulation and excess Na+ extrusion from the cells compared with XJD. The lignin accumulation was higher in the salt-adapted ZM genotype than the sensitive XJD genotype. Consequently, several lignin biosynthesis-related genes including 4CL2, CCoAOMT, COMT, CCR, C4H, PAL1, and PRX1 exhibited higher mRNA expression in salt-tolerant ZM compared with XJD. Moreover, antioxidant enzyme (catalase, superoxide dismutase, ascorbate peroxidase, and glutathione reductase) activity was higher in ZM relative to XJD. This result suggests that high antioxidant provided the defense against oxidative damages in ZM, whereas low enzyme activity with high Na+ triggered the oxidative damage in XJD. These findings together illustrate the ion exchanger, antiporter, and lignin biosysthetic genes involving mechanistic insights into differential salt tolerance in alfalfa.
Keywords: abiotic stress; antioxidant defense; ion exchanger; phenylpropanoid; salinity stress; sodium transporter.
Copyright © 2022 Rahman, Woo, Lee, Park, Kabir, Raza, El Sabagh and Lee.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







References
-
- Bates L. S., Waldren R. P., Teare I. D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39 (1), 205–207. doi: 10.1007/BF00018060 - DOI
-
- Betzen B. M., Smart C. M., Maricle K. L., MariCle B. R. (2019). Effects of increasing salinity on photosynthesis and plant water potential in kansas salt marsh species. Transac. Kansas Acad. Sci. 122(1-2), 49–58, 10. doi: 10.1660/062.122.0105 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous