

Adaptive sequence divergence forged new neurodevelopmental enhancers in humans
- PMID: 36423581
- PMCID: PMC10013929
- DOI: 10.1016/j.cell.2022.10.016
Adaptive sequence divergence forged new neurodevelopmental enhancers in humans
Abstract
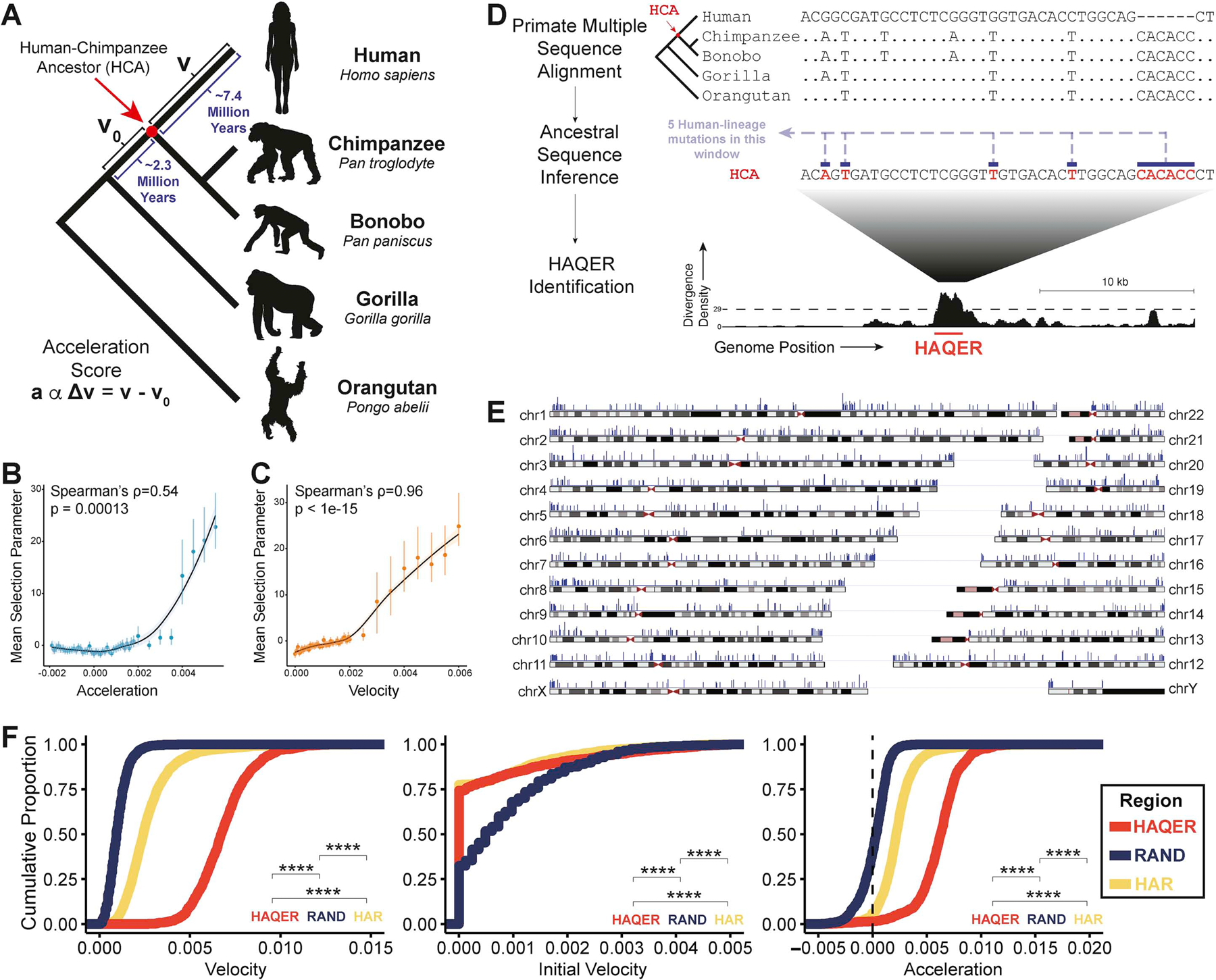
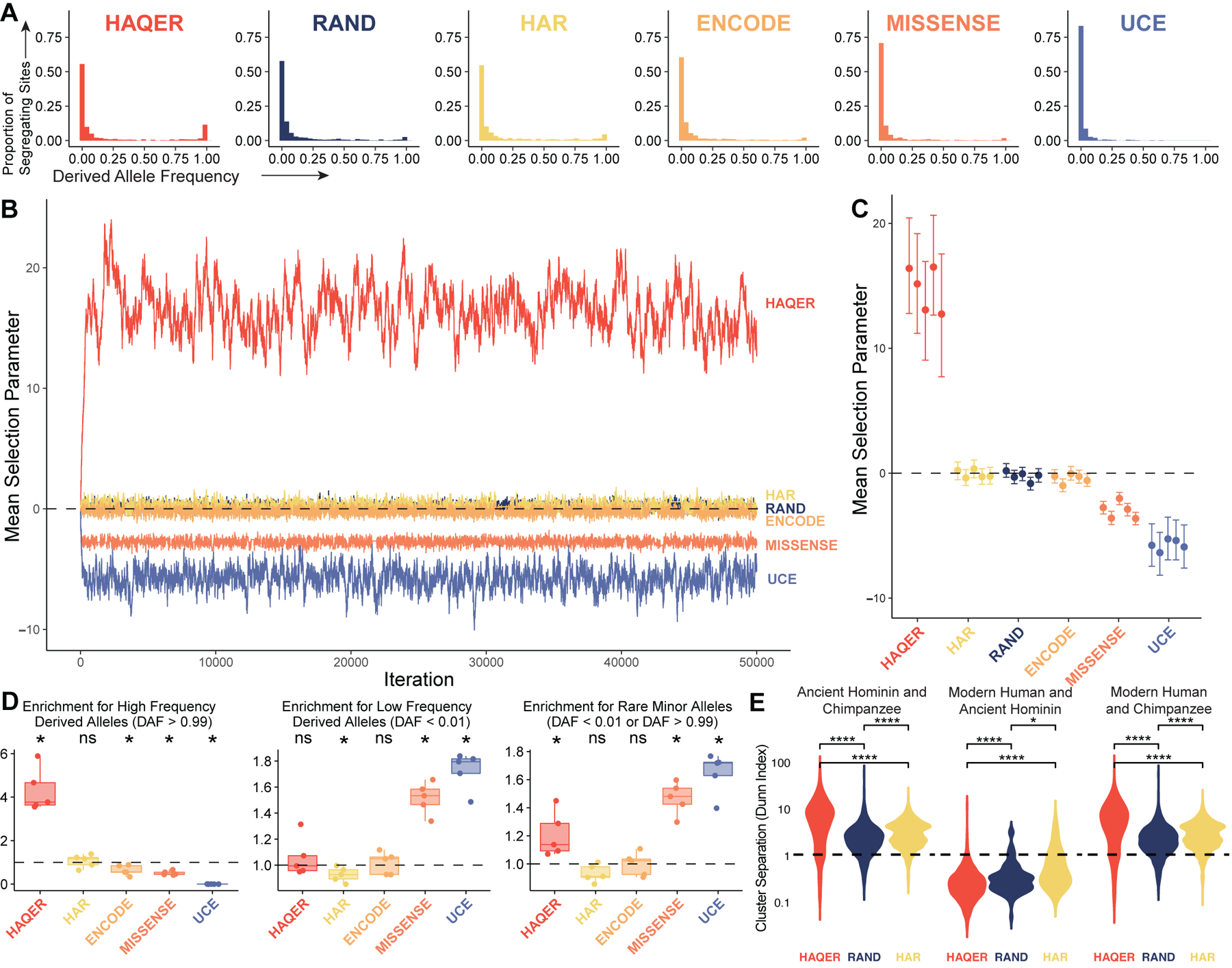
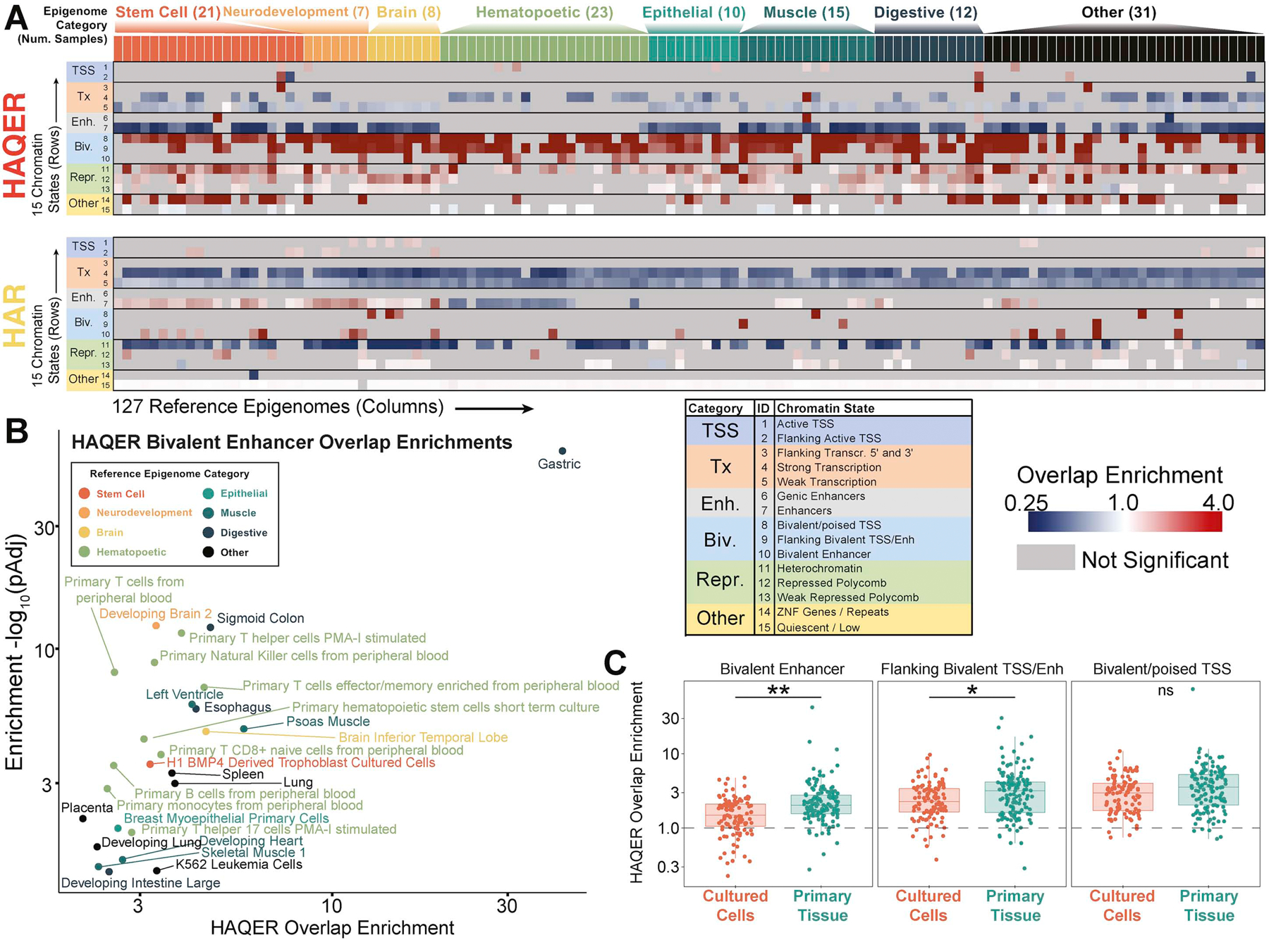
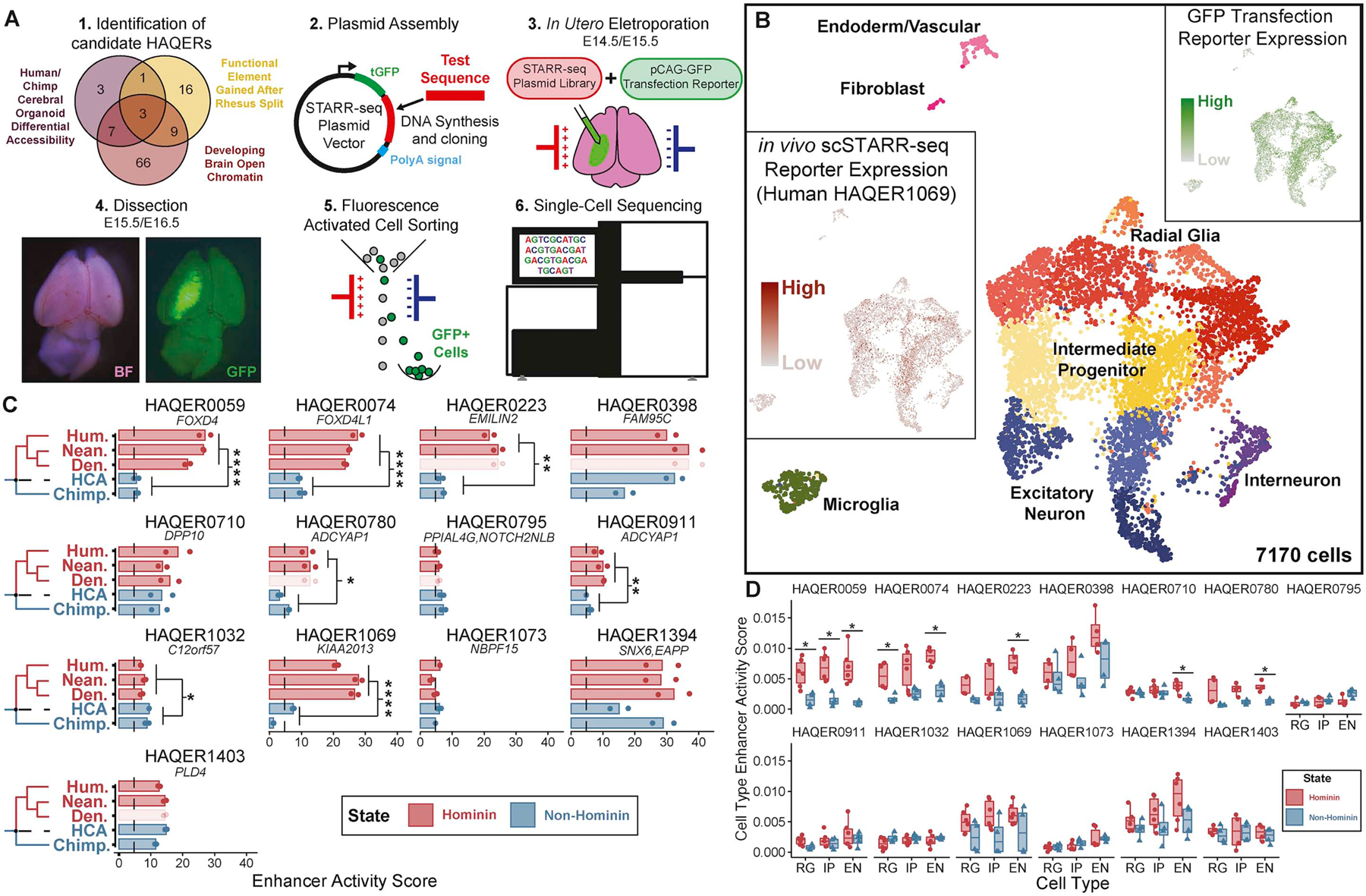
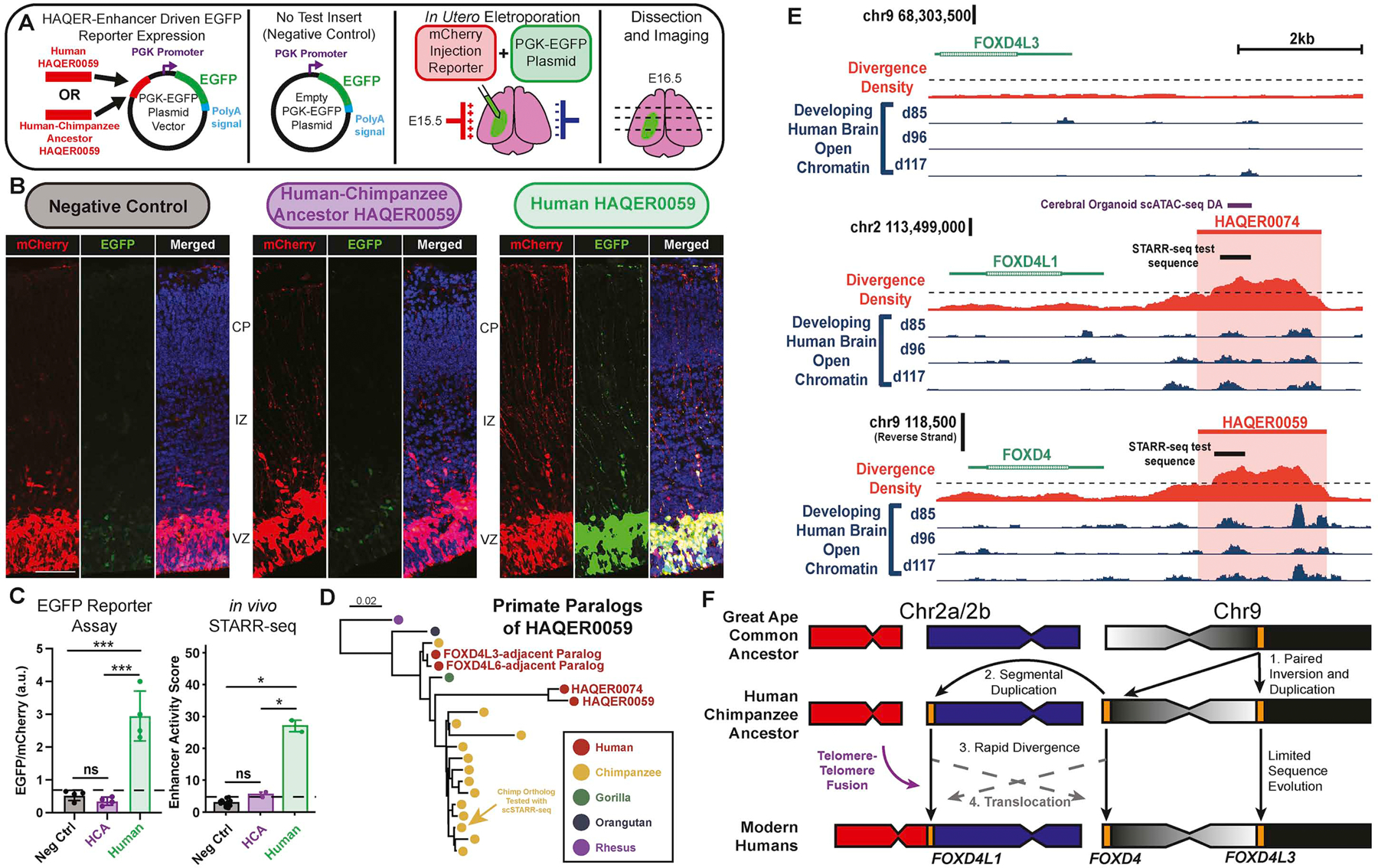
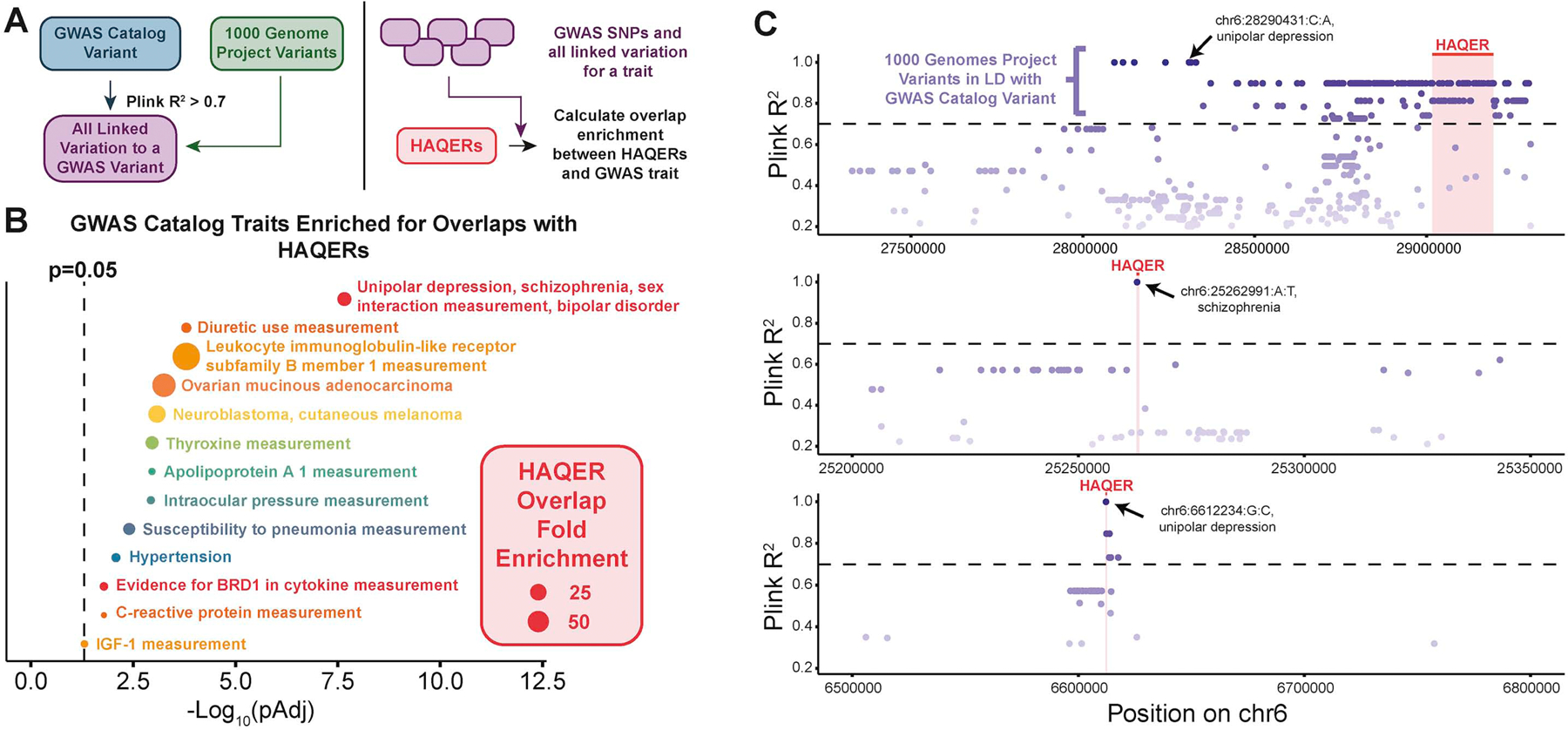
Searches for the genetic underpinnings of uniquely human traits have focused on human-specific divergence in conserved genomic regions, which reflects adaptive modifications of existing functional elements. However, the study of conserved regions excludes functional elements that descended from previously neutral regions. Here, we demonstrate that the fastest-evolved regions of the human genome, which we term "human ancestor quickly evolved regions" (HAQERs), rapidly diverged in an episodic burst of directional positive selection prior to the human-Neanderthal split, before transitioning to constraint within hominins. HAQERs are enriched for bivalent chromatin states, particularly in gastrointestinal and neurodevelopmental tissues, and genetic variants linked to neurodevelopmental disease. We developed a multiplex, single-cell in vivo enhancer assay to discover that rapid sequence divergence in HAQERs generated hominin-unique enhancers in the developing cerebral cortex. We propose that a lack of pleiotropic constraints and elevated mutation rates poised HAQERs for rapid adaptation and subsequent susceptibility to disease.
Keywords: HAQER; comparative genomics; human accelerated regions; human evolution; human genetics; neurodevelopment.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Fast-evolving genomic regions underlie human brain development.Nature. 2023 Feb;614(7946):37-38. doi: 10.1038/d41586-023-00069-2. Nature. 2023. PMID: 36650388 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
