

Fission yeast Dis1 is an unconventional TOG/XMAP215 that induces microtubule catastrophe to drive chromosome pulling
- PMID: 36435910
- PMCID: PMC9701203
- DOI: 10.1038/s42003-022-04271-2
Fission yeast Dis1 is an unconventional TOG/XMAP215 that induces microtubule catastrophe to drive chromosome pulling
Abstract
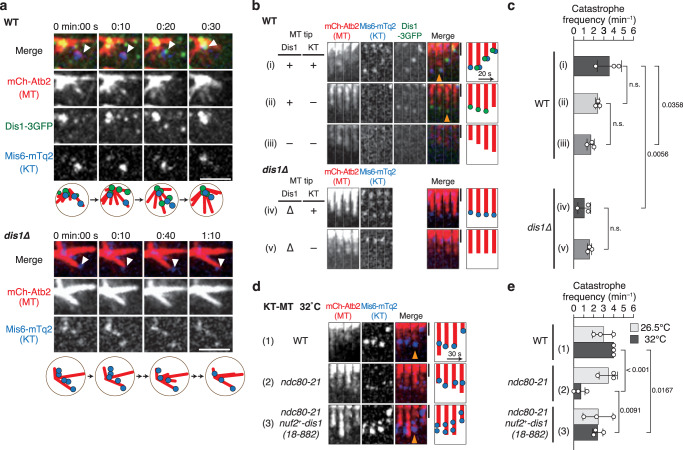
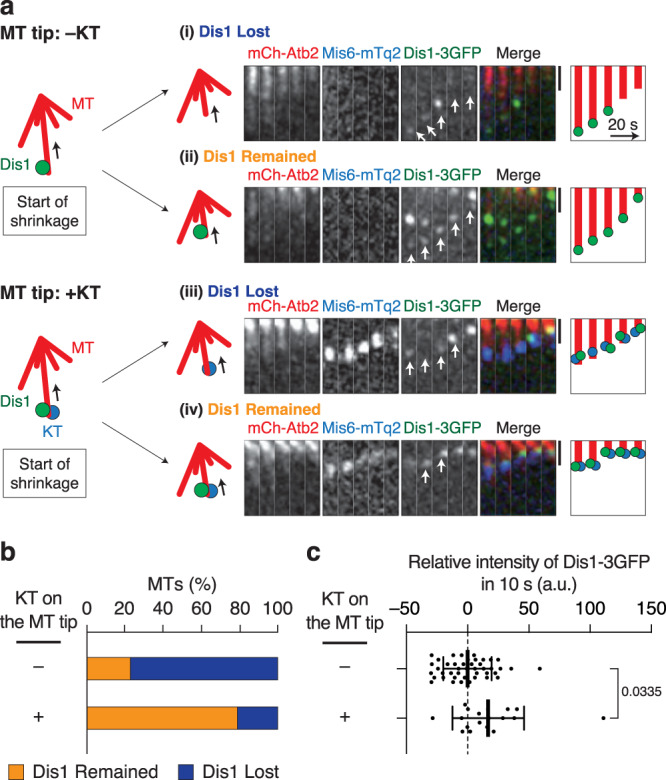
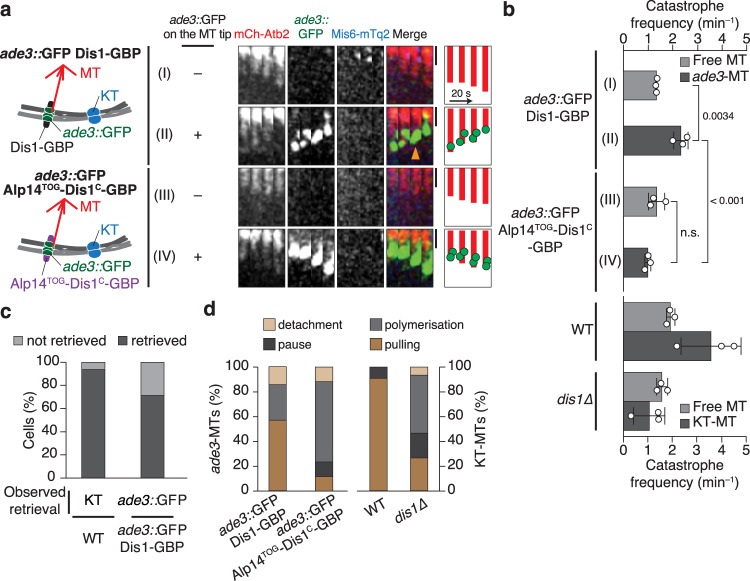
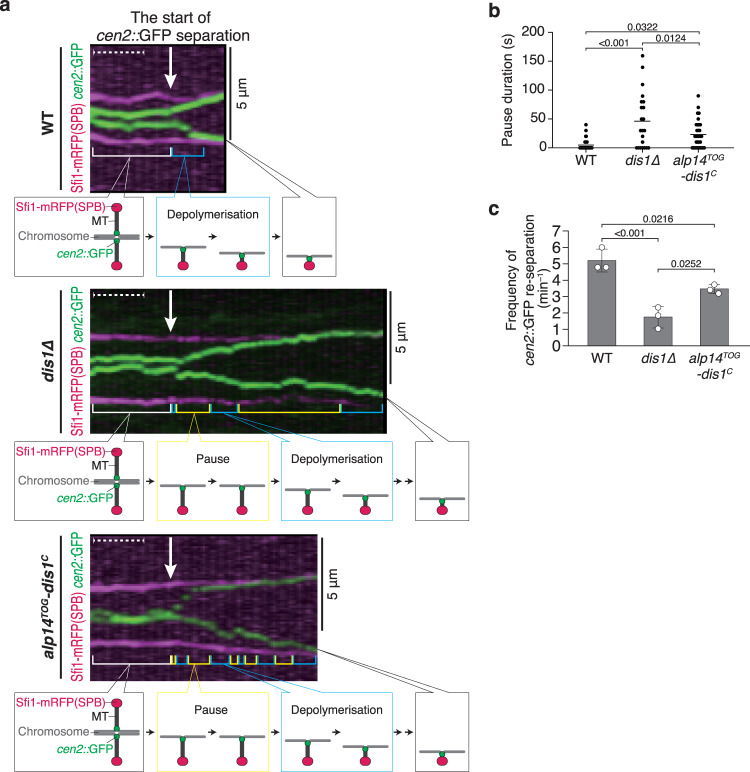
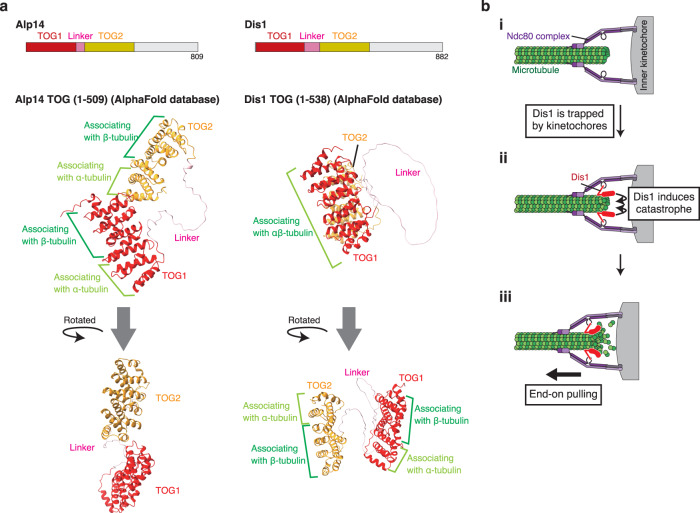
The shortening of microtubules attached to kinetochores is the driving force of chromosome movement during cell division. Specific kinesins are believed to shorten microtubules but are dispensable for viability in yeast, implying the existence of additional factors responsible for microtubule shortening. Here, we demonstrate that Dis1, a TOG/XMAP215 ortholog in fission yeast, promotes microtubule shortening to carry chromosomes. Although TOG/XMAP215 orthologs are generally accepted as microtubule polymerases, Dis1 promoted microtubule catastrophe in vitro and in vivo. Notably, microtubule catastrophe was promoted when the tip was attached to kinetochores, as they steadily anchored Dis1 at the kinetochore-microtubule interface. Engineered Dis1 oligomers artificially tethered at a chromosome arm region induced the shortening of microtubules in contact, frequently pulling the chromosome arm towards spindle poles. This effect was not brought by oligomerised Alp14. Thus, unlike Alp14 and other TOG/XMAP215 orthologs, Dis1 plays an unconventional role in promoting microtubule catastrophe, thereby driving chromosome movement.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Two XMAP215/TOG Microtubule Polymerases, Alp14 and Dis1, Play Non-Exchangeable, Distinct Roles in Microtubule Organisation in Fission Yeast.Int J Mol Sci. 2019 Oct 15;20(20):5108. doi: 10.3390/ijms20205108. Int J Mol Sci. 2019. PMID: 31618856 Free PMC article.
-
Fission yeast ch-TOG/XMAP215 homologue Alp14 connects mitotic spindles with the kinetochore and is a component of the Mad2-dependent spindle checkpoint.EMBO J. 2001 Jul 2;20(13):3389-401. doi: 10.1093/emboj/20.13.3389. EMBO J. 2001. PMID: 11432827 Free PMC article.
-
Microtubules and Alp7-Alp14 (TACC-TOG) reposition chromosomes before meiotic segregation.Nat Cell Biol. 2013 Jul;15(7):786-96. doi: 10.1038/ncb2782. Epub 2013 Jun 16. Nat Cell Biol. 2013. PMID: 23770679
-
MAPping the eukaryotic tree of life: structure, function, and evolution of the MAP215/Dis1 family of microtubule-associated proteins.Int Rev Cytol. 2004;239:179-272. doi: 10.1016/S0074-7696(04)39004-2. Int Rev Cytol. 2004. PMID: 15464854 Review.
-
Regulation of microtubule dynamics by TOG-domain proteins XMAP215/Dis1 and CLASP.Trends Cell Biol. 2011 Oct;21(10):604-14. doi: 10.1016/j.tcb.2011.06.007. Epub 2011 Jul 23. Trends Cell Biol. 2011. PMID: 21782439 Free PMC article. Review.
Cited by
-
Effects of rpl1001 Gene Deletion on Cell Division of Fission Yeast and Its Molecular Mechanism.Curr Issues Mol Biol. 2024 Mar 18;46(3):2576-2597. doi: 10.3390/cimb46030164. Curr Issues Mol Biol. 2024. PMID: 38534780 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
