

Early microbial exposure shapes adult immunity by altering CD8+ T cell development
- PMID: 36442114
- PMCID: PMC9894172
- DOI: 10.1073/pnas.2212548119
Early microbial exposure shapes adult immunity by altering CD8+ T cell development
Abstract
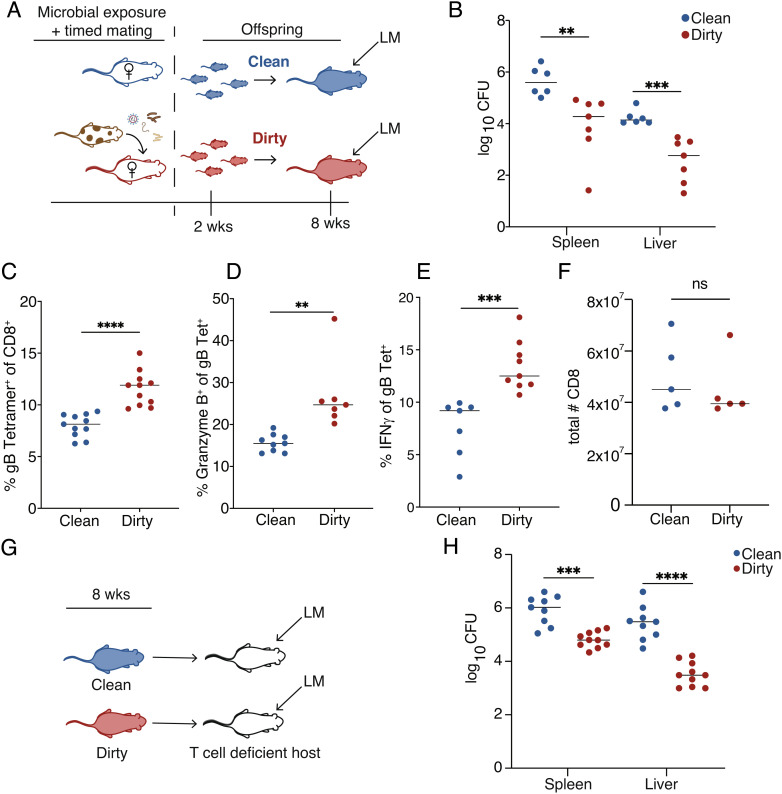
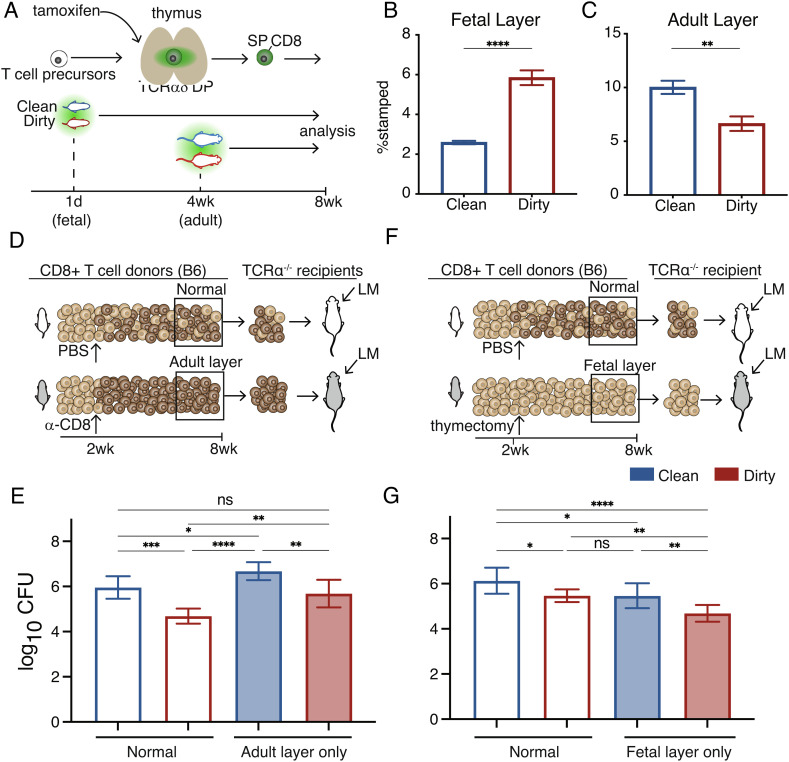
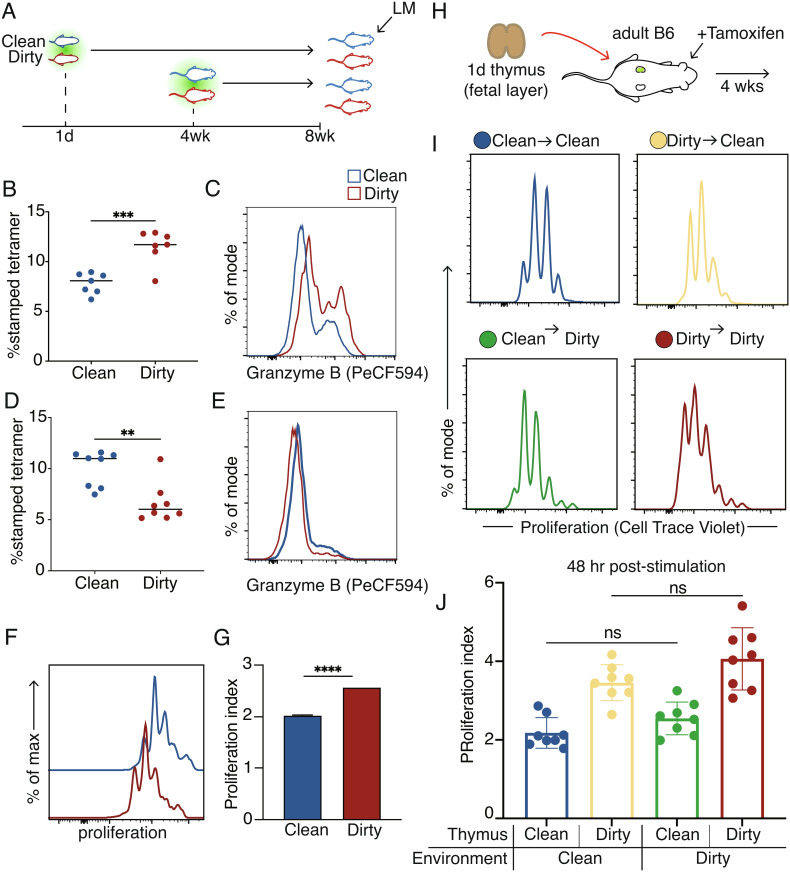
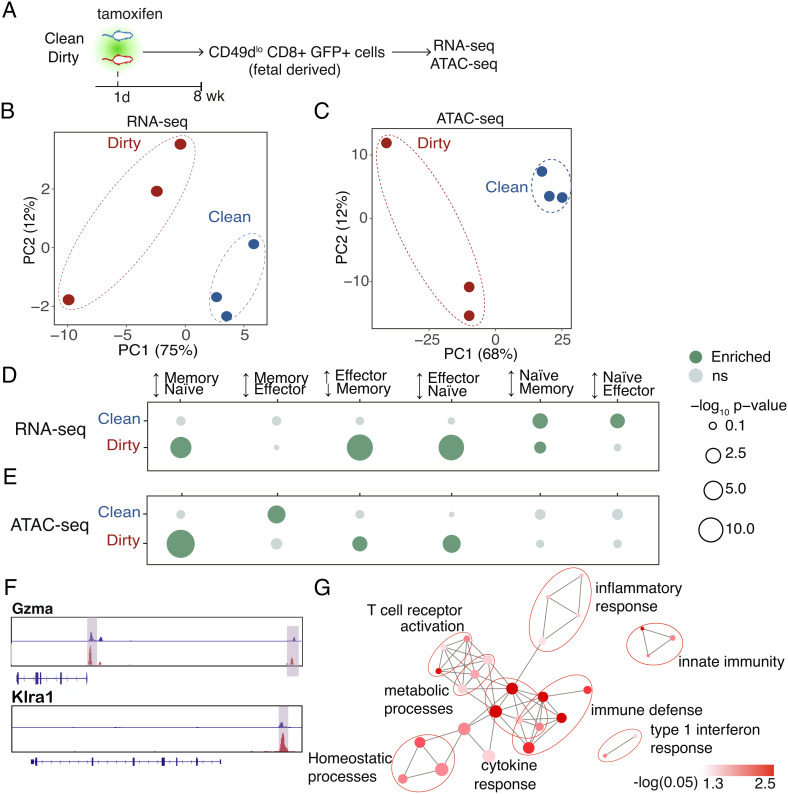
Microbial exposure during development can elicit long-lasting effects on the health of an individual. However, how microbial exposure in early life leads to permanent changes in the immune system is unknown. Here, we show that the microbial environment alters the set point for immune susceptibility by altering the developmental architecture of the CD8+ T cell compartment. In particular, early microbial exposure results in the preferential expansion of highly responsive fetal-derived CD8+ T cells that persist into adulthood and provide the host with enhanced immune protection against intracellular pathogens. Interestingly, microbial education of fetal-derived CD8+ T cells occurs during thymic development rather than in the periphery and involves the acquisition of a more effector-like epigenetic program. Collectively, our results provide a conceptual framework for understanding how microbial colonization in early life leads to lifelong changes in the immune system.
Keywords: CD8+ T cells; developmental layering; developmental origins of adult health and disease; immune training; pet-shop dirty mouse model.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
