

Loss of FoxOs in muscle increases strength and mitochondrial function during aging
- PMID: 36442857
- PMCID: PMC9891940
- DOI: 10.1002/jcsm.13124
Loss of FoxOs in muscle increases strength and mitochondrial function during aging
Abstract
Background: Muscle mitochondrial decline is associated with aging-related muscle weakness and insulin resistance. FoxO transcription factors are targets of insulin action and deletion of FoxOs improves mitochondrial function in diabetes. However, disruptions in proteostasis and autophagy are hallmarks of aging and the effect of chronic inhibition of FoxOs in aged muscle is unknown. This study investigated the role of FoxOs in regulating muscle strength and mitochondrial function with age.
Methods: We measured muscle strength, cross-sectional area, muscle fibre-type, markers of protein synthesis/degradation, central nuclei, glucose/insulin tolerance, and mitochondrial bioenergetics in 4.5-month (Young) and 22-24-month-old (Aged) muscle-specific FoxO1/3/4 triple KO (TKO) and littermate control (Ctrl) mice.
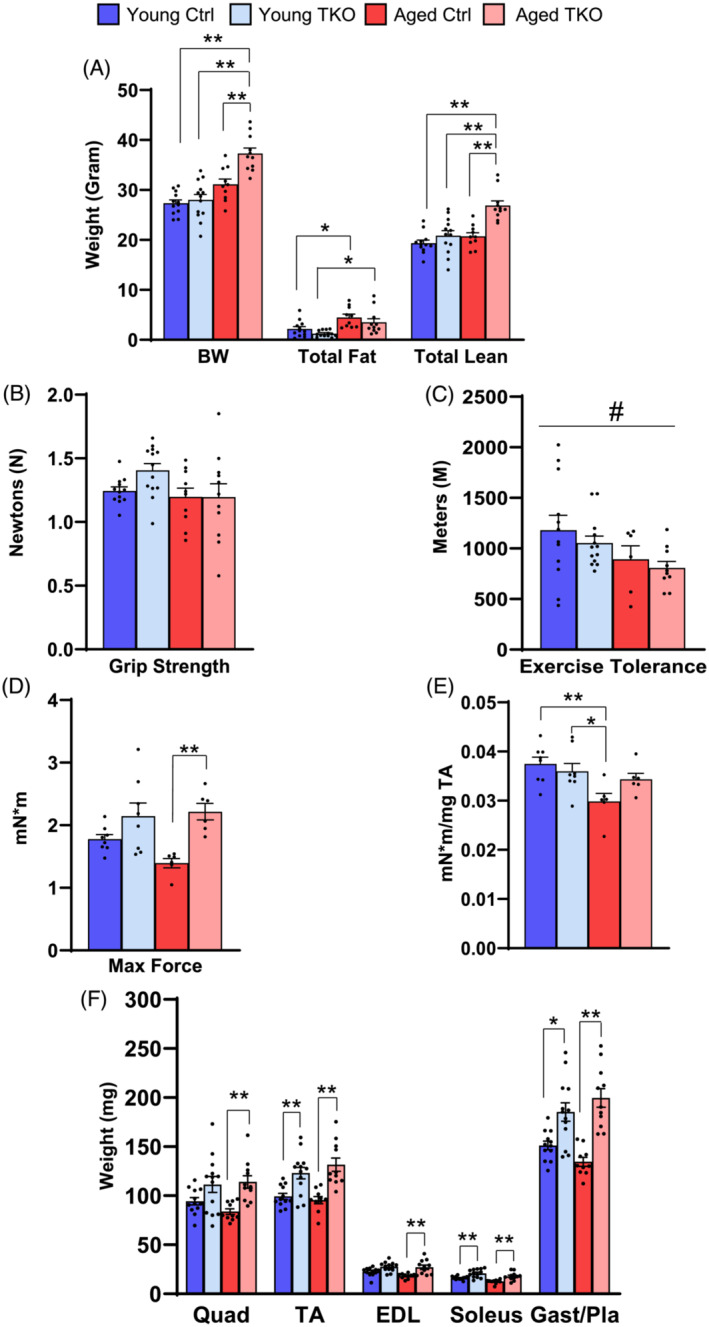
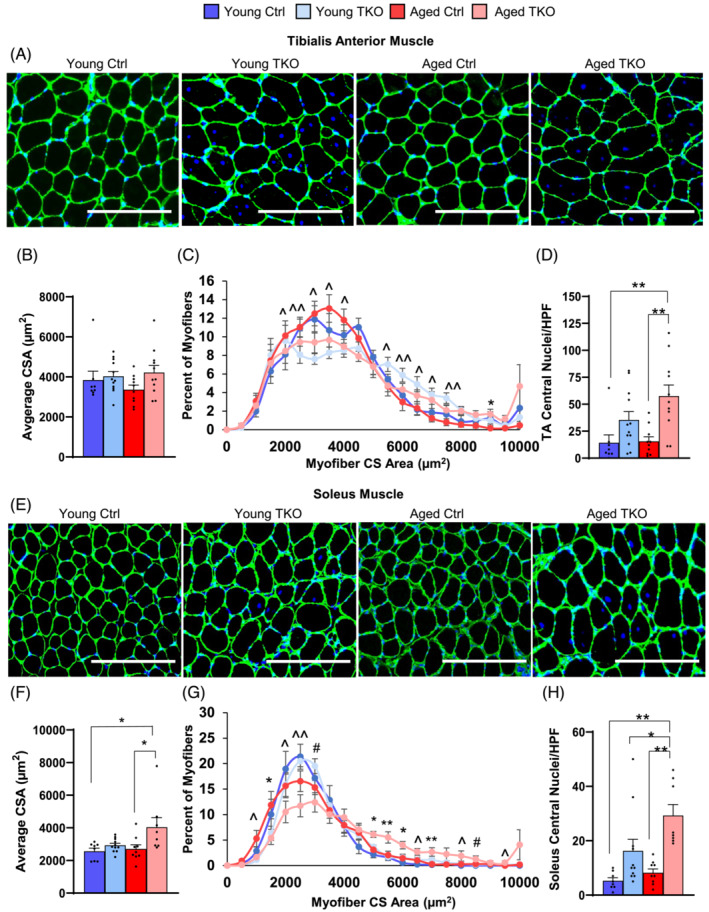
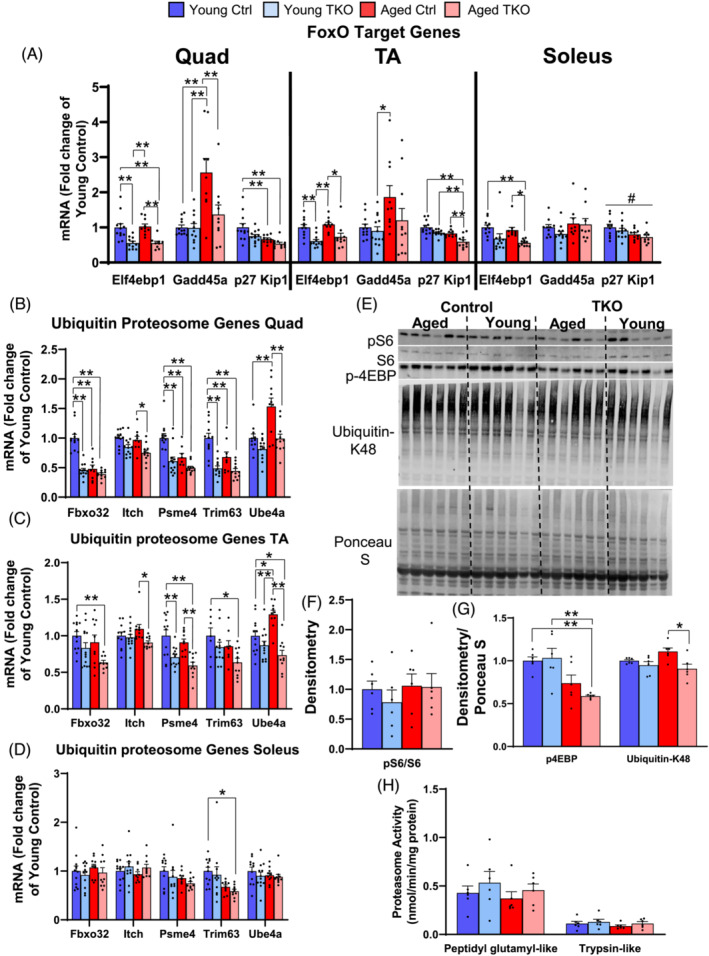
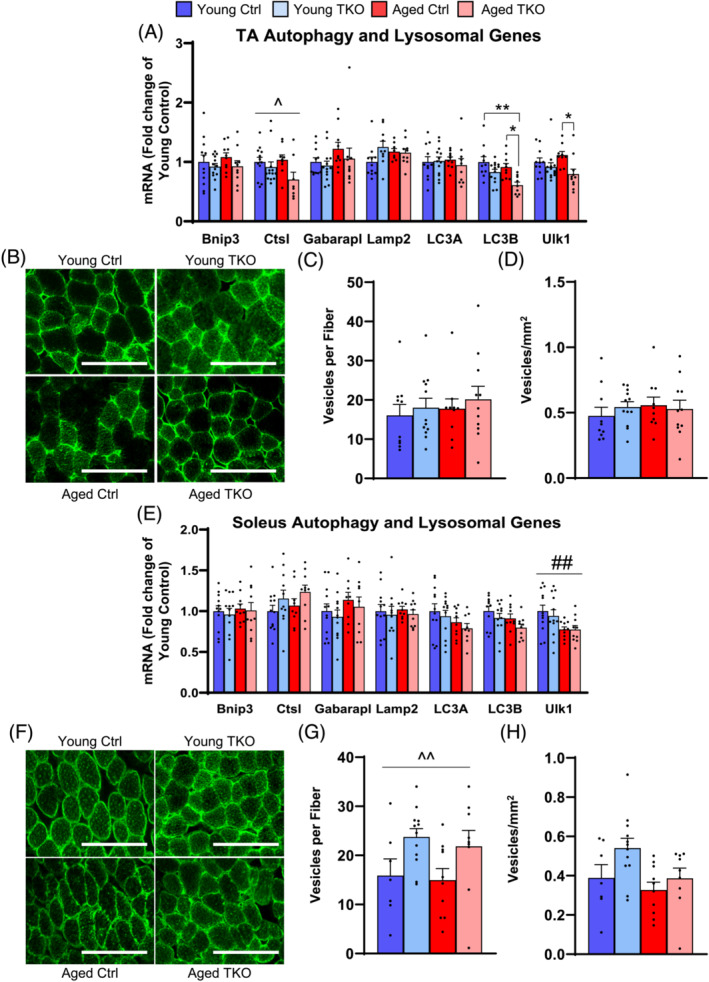
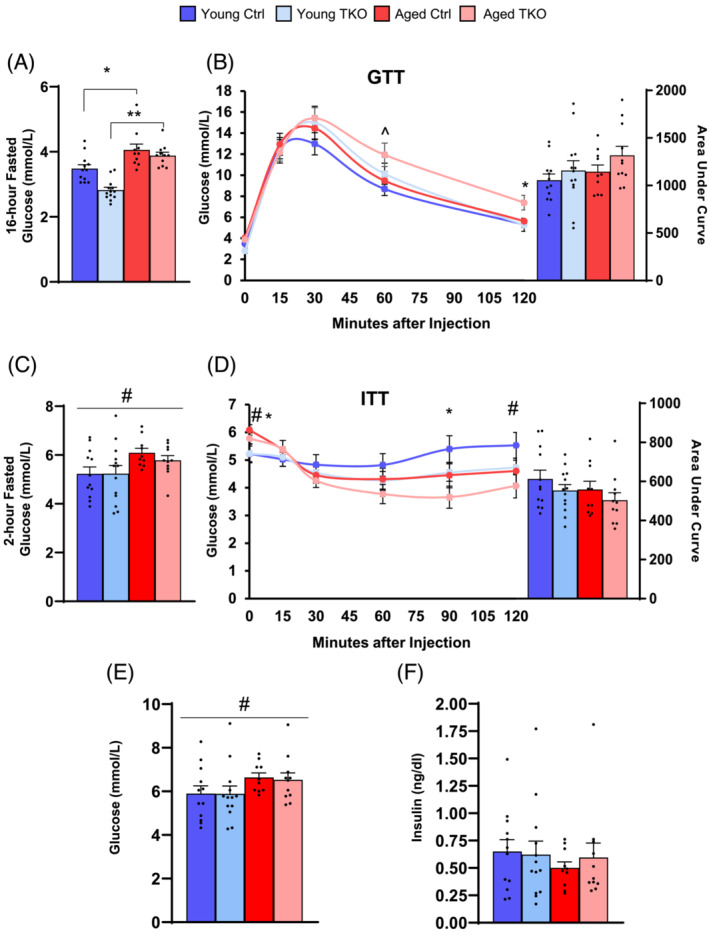
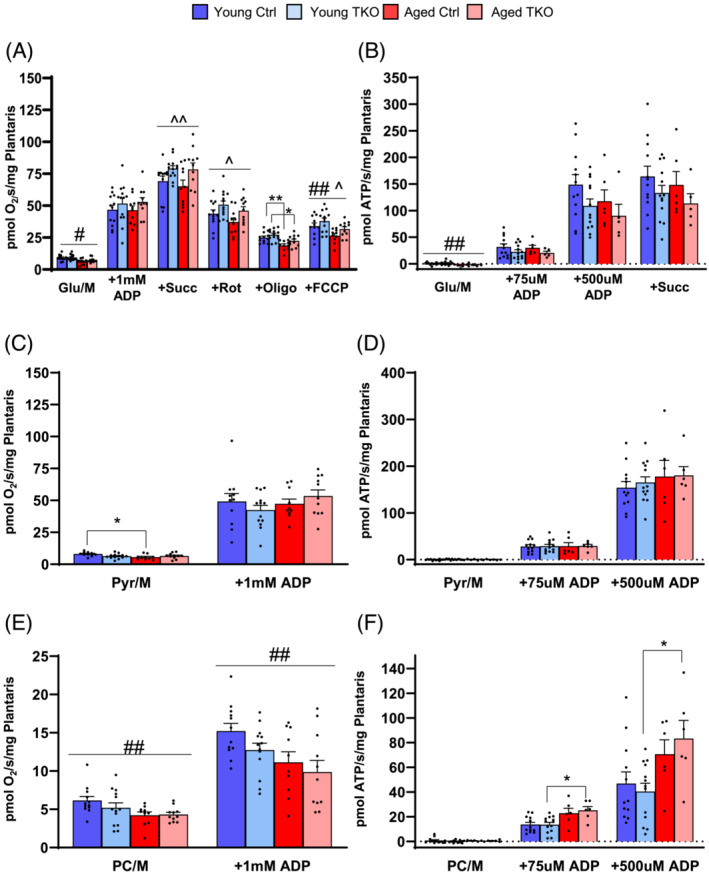
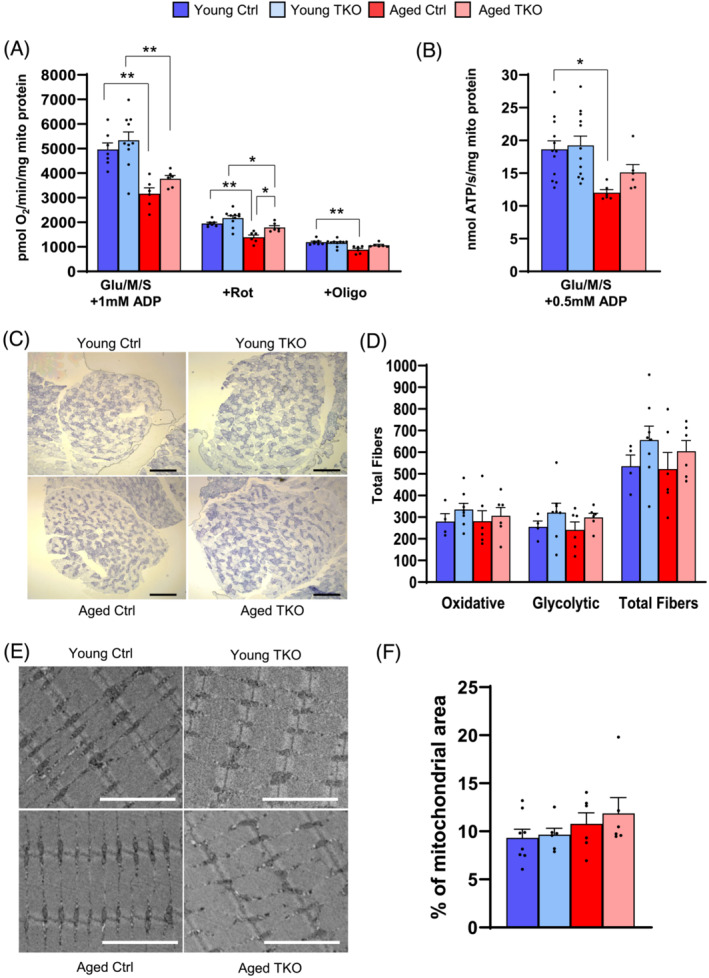
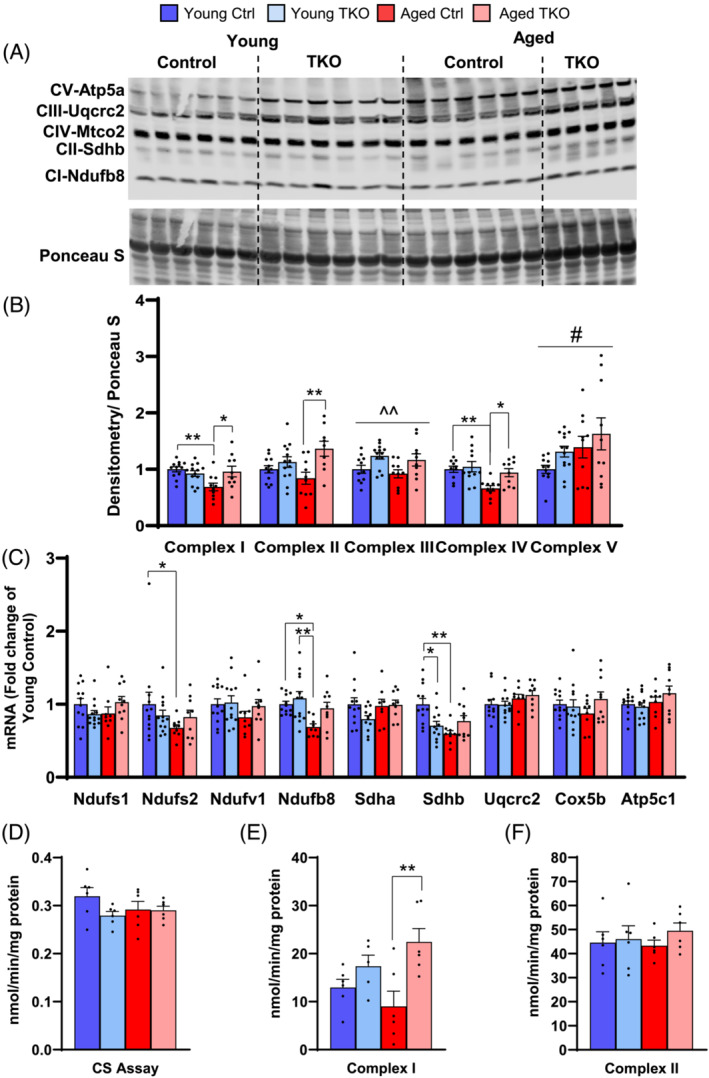
Results: Lean mass was increased in Aged TKO compared with both Aged Ctrl and younger groups by 26-33% (P < 0.01). Muscle strength, measured by max force of tibialis anterior (TA) contraction, was 20% lower in Aged Ctrl compared with Young Ctrls (P < 0.01) but was not decreased in Aged TKOs. Increased muscle strength in Young and Aged TKO was associated with 18-48% increased muscle weights compared with Ctrls (P < 0.01). Muscle cross-sectional analysis of TA, soleus, and plantaris revealed increases in fibre size distribution and a 2.5-10-fold increase in central nuclei in Young and Aged TKO mice, without histologic signs of muscle damage. Age-dependent increases in Gadd45a and Ube4a expression as well accumulation of K48 polyubiquitinated proteins were observed in quad and TA but were prevented by FoxO deletion. Young and Aged TKO muscle showed minimal changes in autophagy flux and no accumulation of autophagosomes compared with Ctrl groups. Increased strength in Young and Aged TKO was associated with a 10-20% increase in muscle mitochondrial respiration using glutamate/malate/succinate compared with controls (P < 0.05). OXPHOS subunit expression and complex I activity were decreased 16-34% in Aged Ctrl compared with Young Ctrl but were prevented in Aged TKO. Both Aged Ctrl and Aged TKO showed impaired glucose tolerance by 33% compared to young groups (P < 0.05) indicating improved strength and mitochondrial respiration are not due to improved glycemia.
Conclusions: FoxO deletion increases muscle strength even during aging. Deletion of FoxOs maintains muscle strength in part by mild suppression of atrophic pathways, including inhibition of Gadd45a and Ube4a expression, without accumulation of autophagosomes in muscle. Deletion of FoxOs also improved mitochondrial function by maintenance of OXPHOS in both young and aged TKO.
Keywords: Aging; FoxO; Glucose tolerance; Insulin resistance; Mitochondrial function; Muscle hypertrophy.
© 2022 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
The authors declare no conflict of interest regarding the present work.
Figures
References
-
- Goodpaster BH, Park SW, Harris TB, Kritchevsky SB, Nevitt M, Schwartz AV, et al. The loss of skeletal muscle strength, mass, and quality in older adults: the health, aging and body composition study. J Gerontol A Biol Sci Med Sci 2006;61:1059–1064. - PubMed
-
- Newman AB, Kupelian V, Visser M, Simonsick EM, Goodpaster BH, Kritchevsky SB, et al. Strength, but not muscle mass, is associated with mortality in the health, aging and body composition study cohort. J Gerontol A Biol Sci Med Sci 2006;61:72–77. - PubMed
-
- Morton RW, Traylor DA, Weijs PJM, Phillips SM. Defining anabolic resistance: implications for delivery of clinical care nutrition. Curr Opin Crit Care 2018;24:124–130. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
