

Non-canonical Wnt signaling in the eye
- PMID: 36443219
- PMCID: PMC10209355
- DOI: 10.1016/j.preteyeres.2022.101149
Non-canonical Wnt signaling in the eye
Abstract
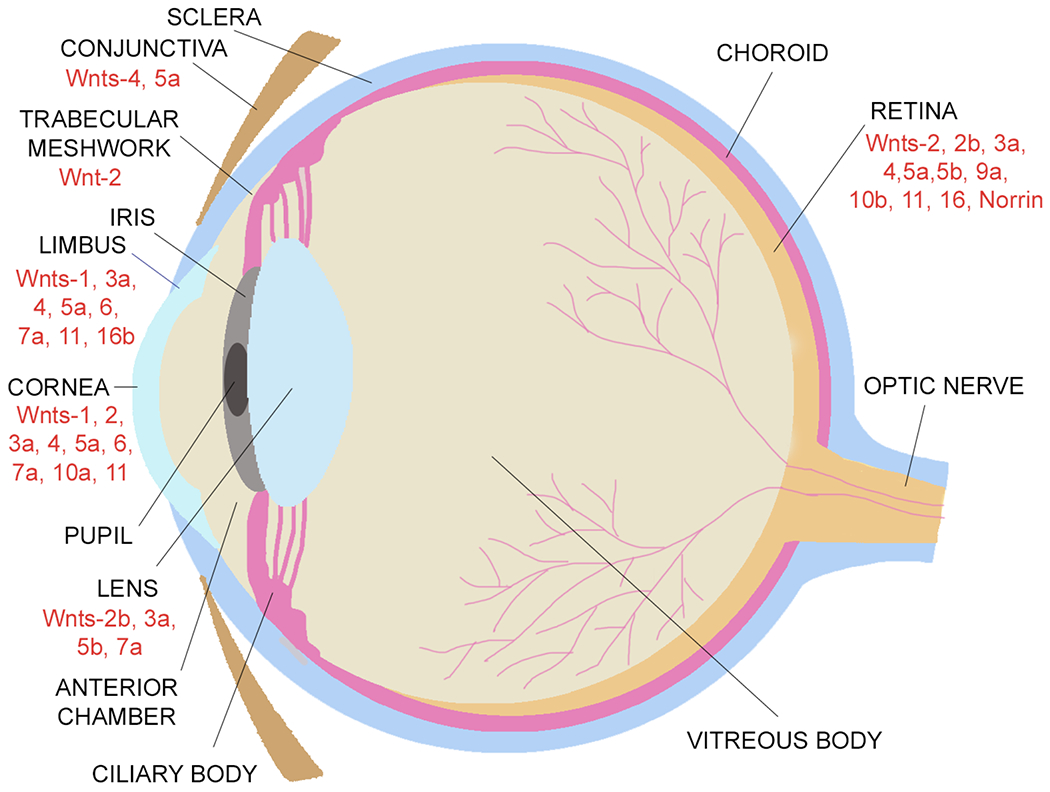
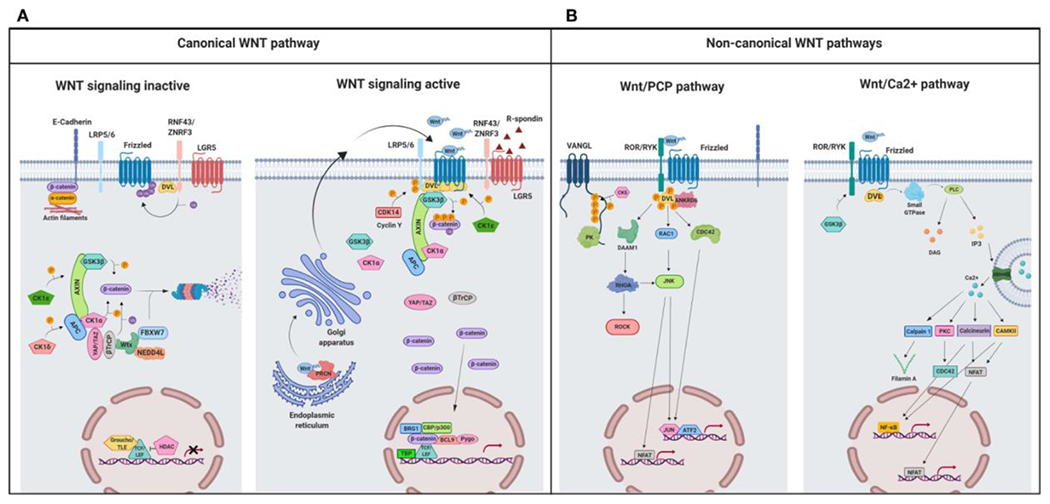
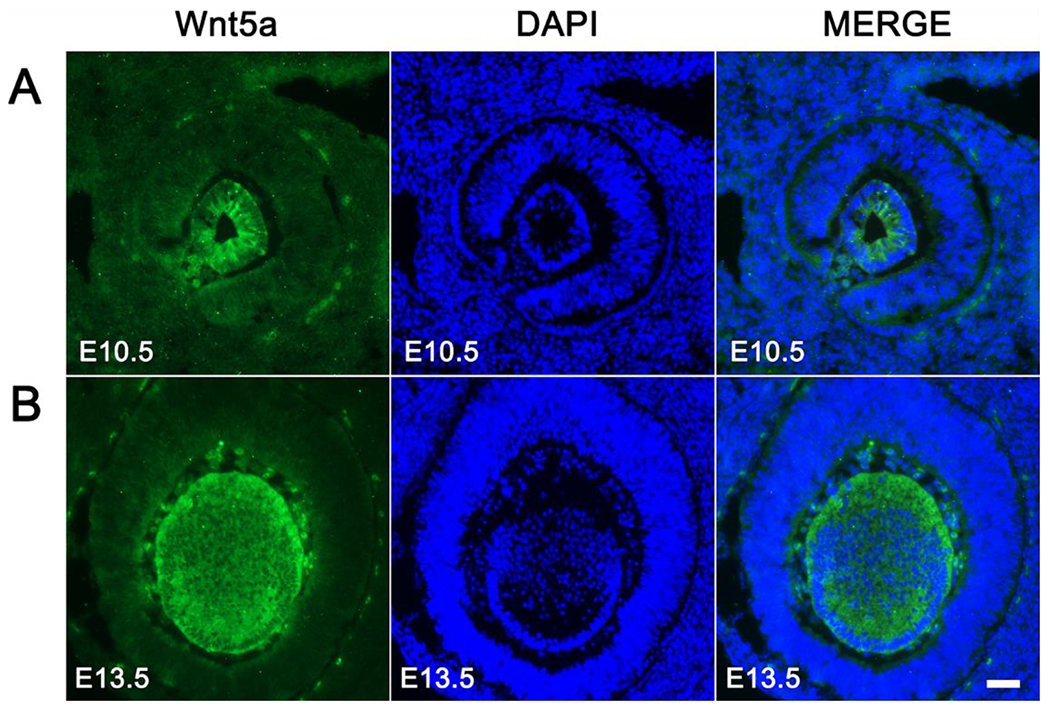
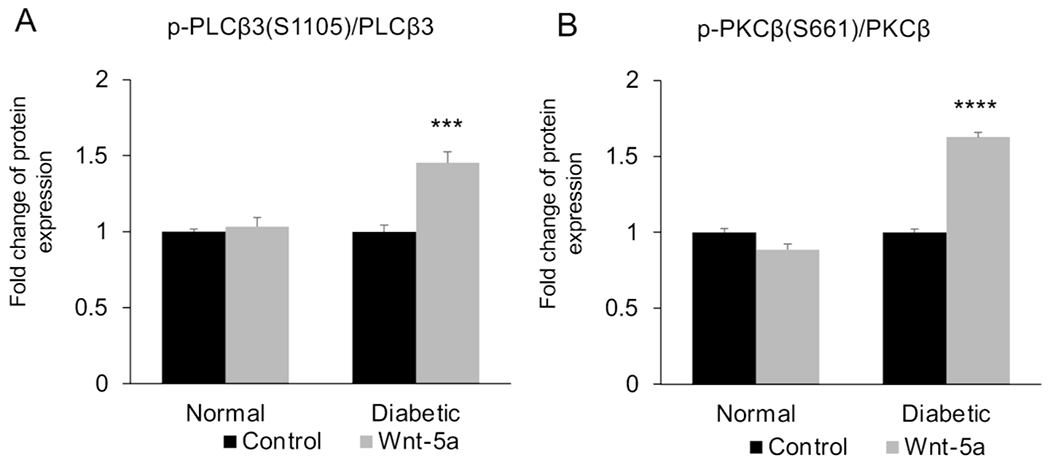
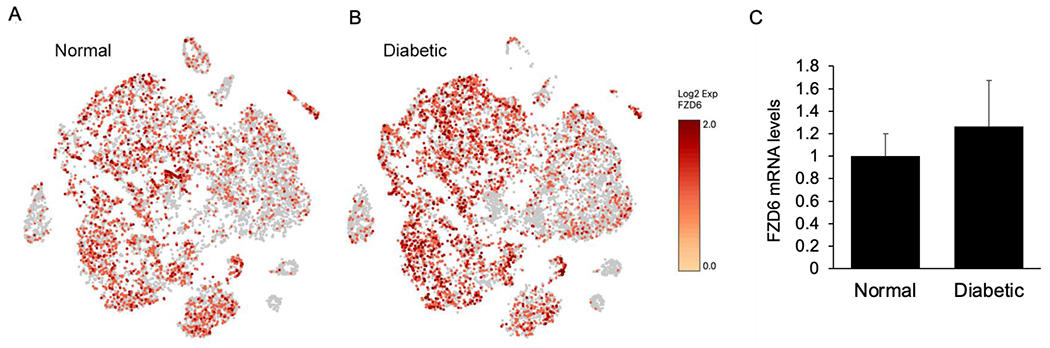
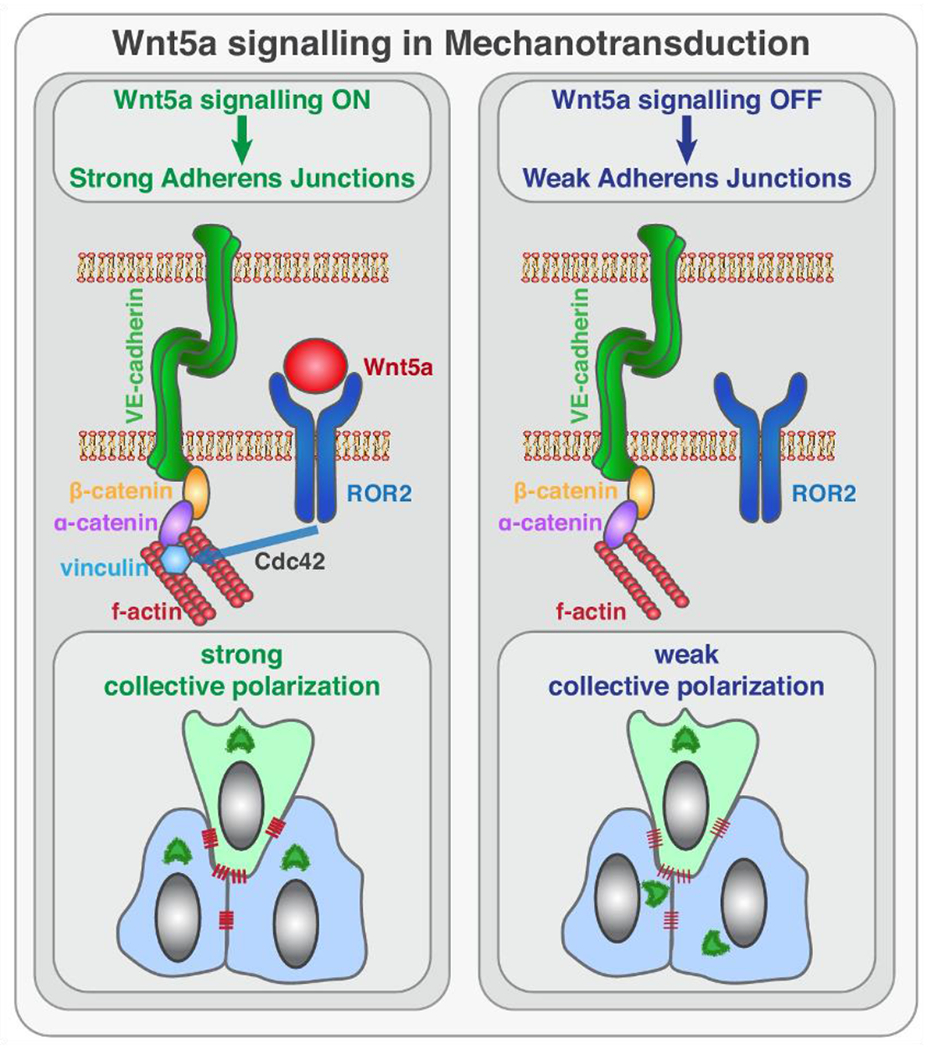
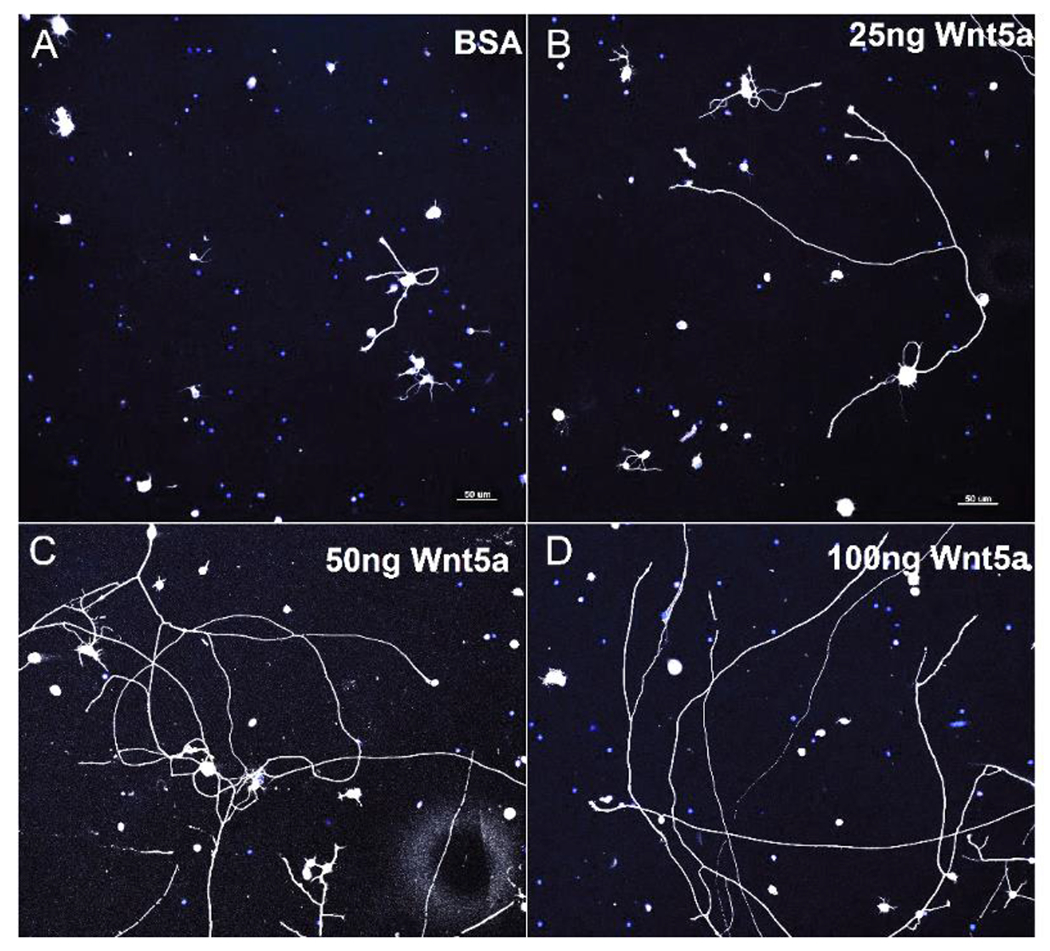
Wnt signaling comprises a group of complex signal transduction pathways that play critical roles in cell proliferation, differentiation, and apoptosis during development, as well as in stem cell maintenance and adult tissue homeostasis. Wnt pathways are classified into two major groups, canonical (β-catenin-dependent) or non-canonical (β-catenin-independent). Most previous studies in the eye have focused on canonical Wnt signaling, and the role of non-canonical signaling remains poorly understood. Additionally, the crosstalk between canonical and non-canonical Wnt signaling in the eye has hardly been explored. In this review, we present an overview of available data on ocular non-canonical Wnt signaling, including developmental and functional aspects in different eye compartments. We also discuss important changes of this signaling in various ocular conditions, such as keratoconus, aniridia-related keratopathy, diabetes, age-related macular degeneration, optic nerve damage, pathological angiogenesis, and abnormalities in the trabecular meshwork and conjunctival cells, and limbal stem cell deficiency.
Keywords: Calcium signaling; Conjunctiva; Cornea; Eye development; Eye diseases; Lens; Planar cell polarity; Retina; Stem cells; Wnt signaling.
Copyright © 2022. Published by Elsevier Ltd.
Conflict of interest statement
Declaration of competing interest None.
Figures
References
-
- Asimellis G, Kaufman EJ. Keratoconus. [Updated 2022 May 30]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2022. Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK470435/
