Active DNA damage response signaling initiates and maintains meiotic sex chromosome inactivation
- PMID: 36443288
- PMCID: PMC9705562
- DOI: 10.1038/s41467-022-34295-5
Active DNA damage response signaling initiates and maintains meiotic sex chromosome inactivation
Abstract
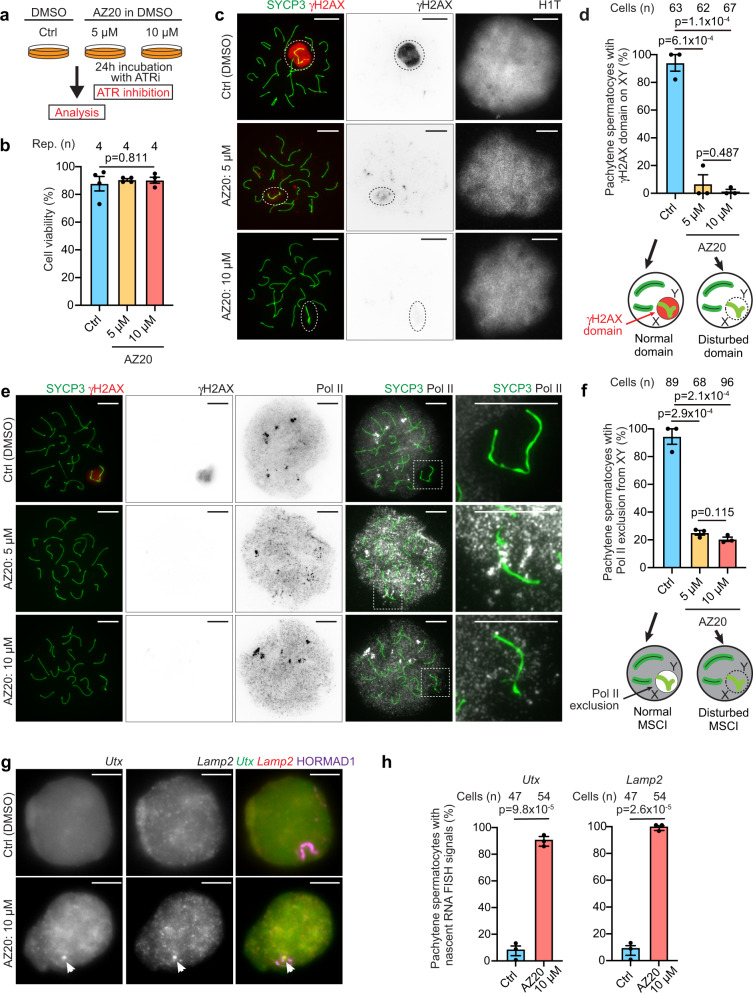
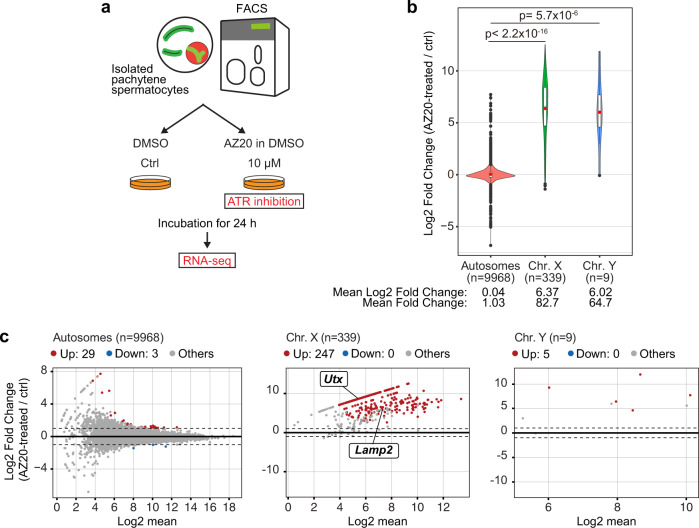
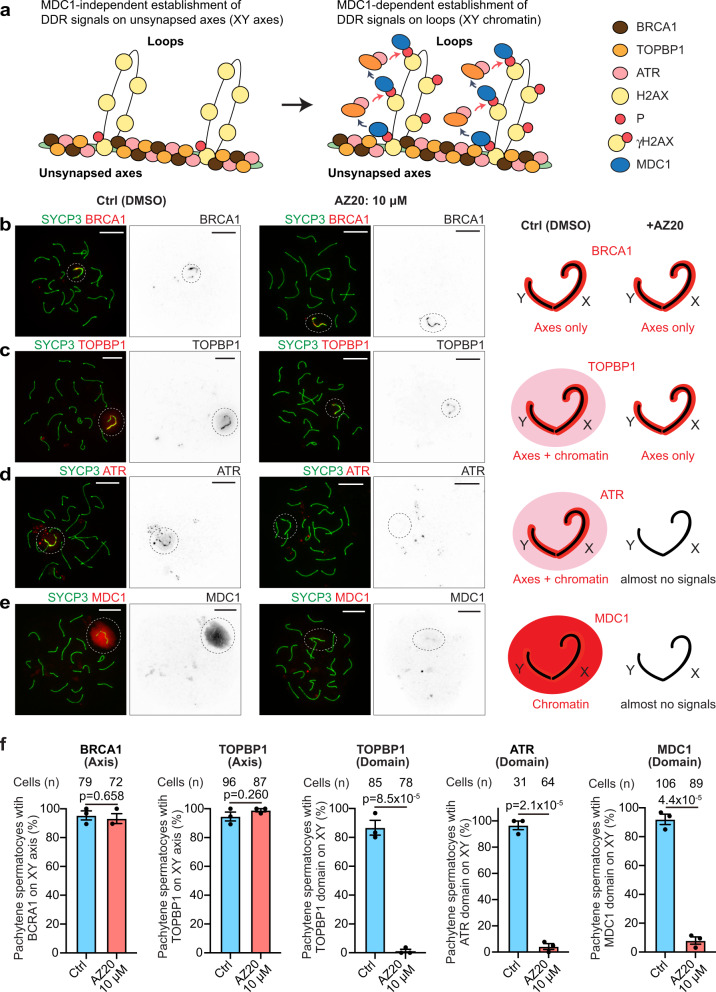
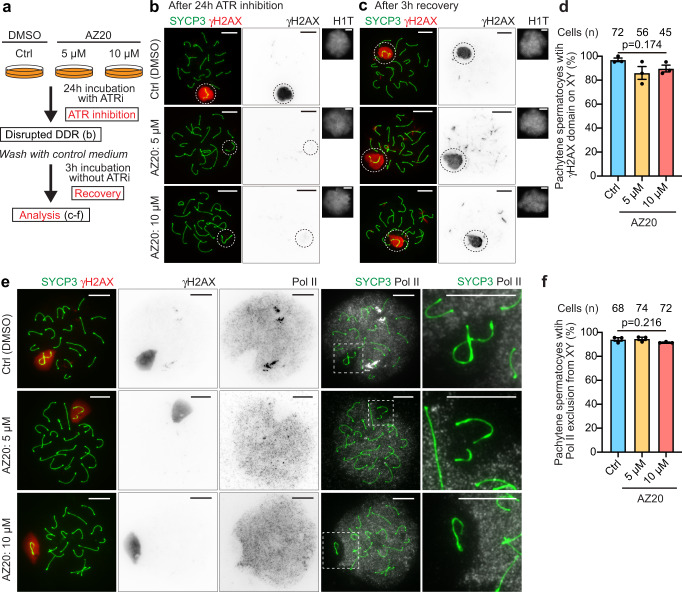
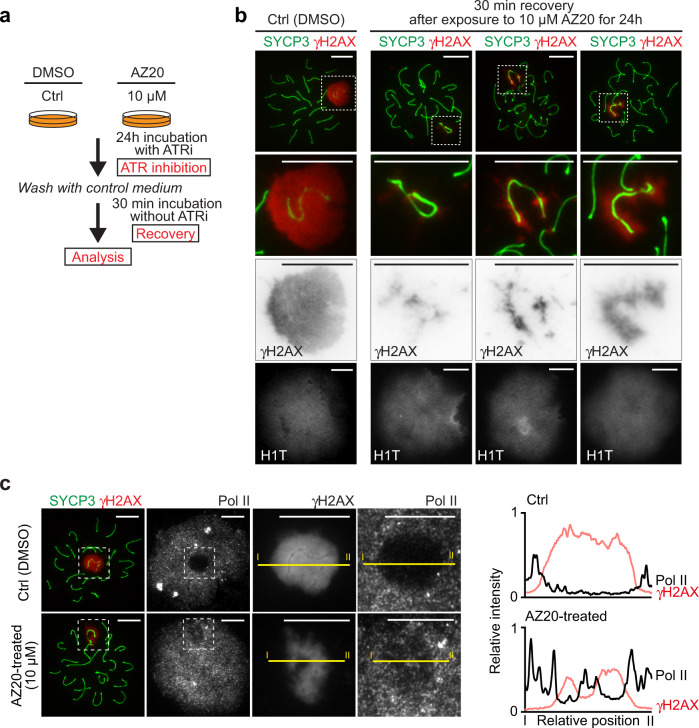
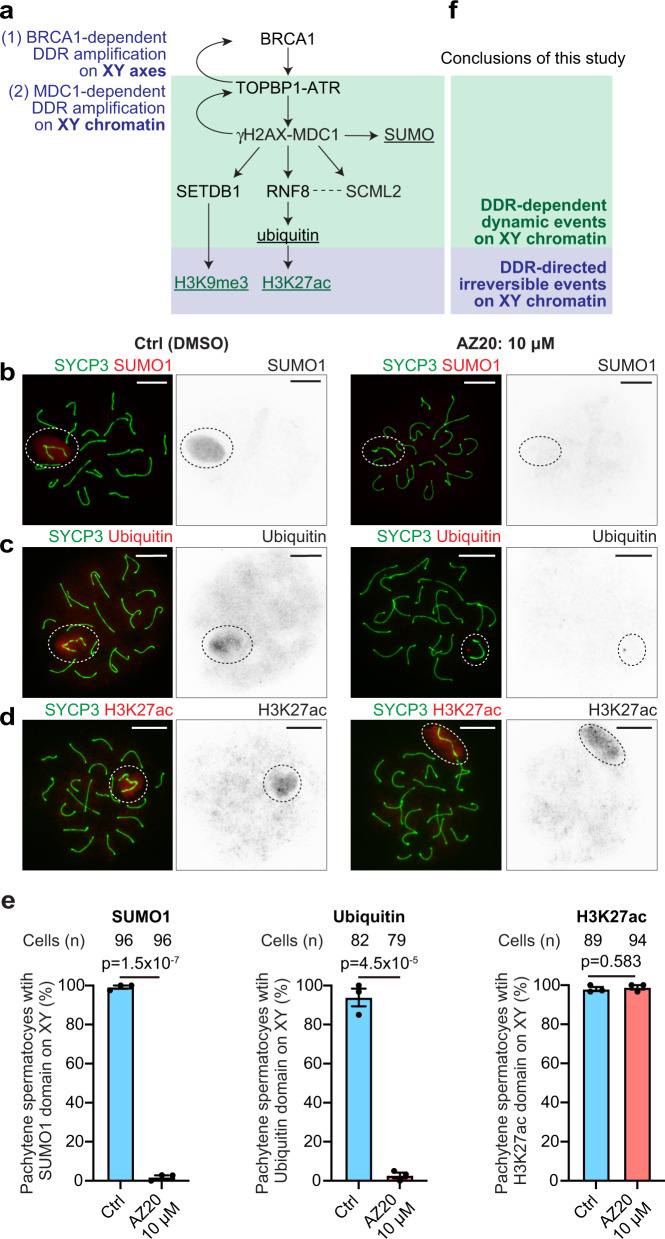
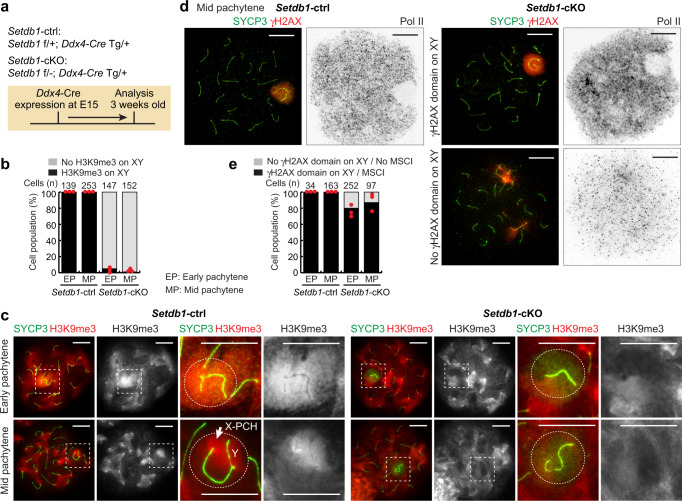
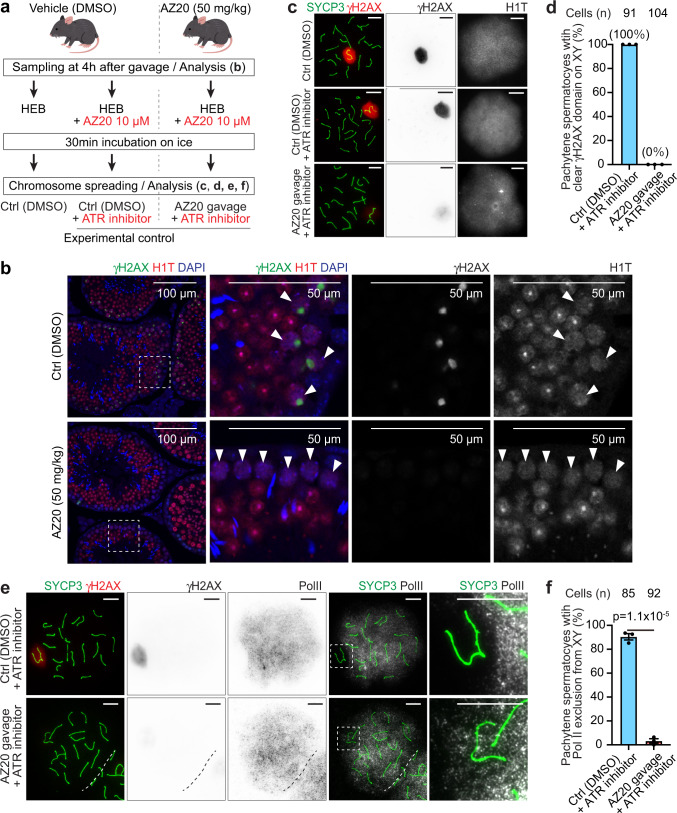
Meiotic sex chromosome inactivation (MSCI) is an essential process in the male germline. While genetic experiments have established that the DNA damage response (DDR) pathway directs MSCI, due to limitations to the experimental systems available, mechanisms underlying MSCI remain largely unknown. Here we establish a system to study MSCI ex vivo, based on a short-term culture method, and demonstrate that active DDR signaling is required both to initiate and maintain MSCI via a dynamic and reversible process. DDR-directed MSCI follows two layers of modifications: active DDR-dependent reversible processes and irreversible histone post-translational modifications. Further, the DDR initiates MSCI independent of the downstream repressive histone mark H3K9 trimethylation (H3K9me3), thereby demonstrating that active DDR signaling is the primary mechanism of silencing in MSCI. By unveiling the dynamic nature of MSCI, and its governance by active DDR signals, our study highlights the sex chromosomes as an active signaling hub in meiosis.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures