

B Cell Activation Results in IKK-Dependent, but Not c-Rel- or RelA-Dependent, Decreases in Transcription of the B Cell Tolerance-Inducing Gene Ets1
- PMID: 36445360
- PMCID: PMC10069408
- DOI: 10.4049/immunohorizons.2100065
B Cell Activation Results in IKK-Dependent, but Not c-Rel- or RelA-Dependent, Decreases in Transcription of the B Cell Tolerance-Inducing Gene Ets1
Abstract
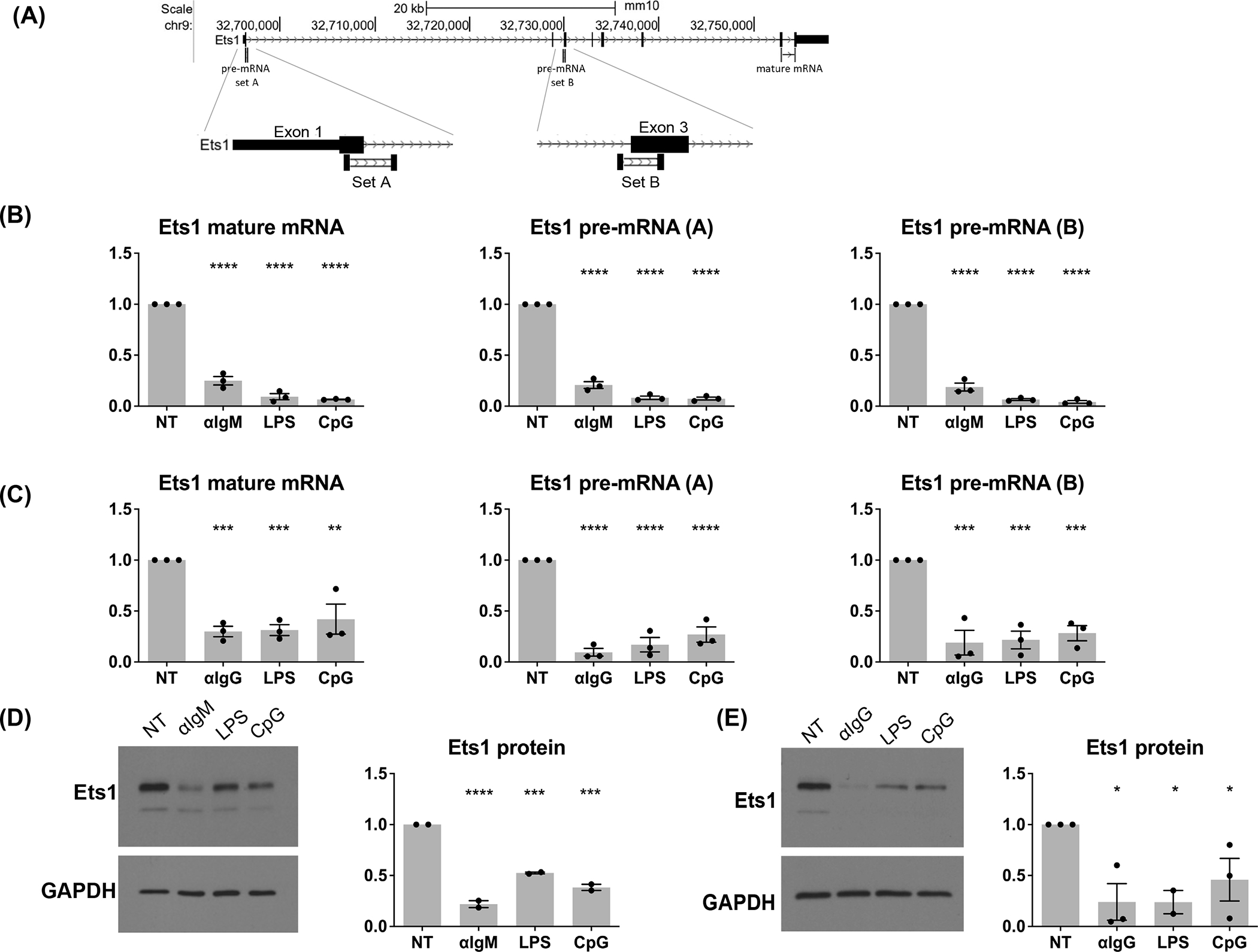
Ets1 is a key transcription factor in B cells that is required to prevent premature differentiation into Ab-secreting cells. Previously, we showed that BCR and TLR signaling downregulate Ets1 levels and that the kinases PI3K, Btk, IKK, and JNK are required for this process. PI3K is important in activating Btk by generating the membrane lipid phosphatidylinositol (3,4,5)-trisphosphate, to which Btk binds via its PH domain. Btk in turn is important in activating the IKK kinase pathway, which it does by activating phospholipase Cγ2→protein kinase Cβ signaling. In this study, we have further investigated the pathways regulating Ets1 in mouse B cells. Although IKK is well known for its role in activating the canonical NF-κB pathway, IKK-mediated downregulation of Ets1 does not require either RelA or c-Rel. We also examined the potential roles of two other IKK targets that are not part of the NF-κB signaling pathway, Foxo3a and mTORC2, in regulating Ets1. We find that loss of Foxo3a or inhibition of mTORC2 does not block BCR-induced Ets1 downregulation. Therefore, these two pathways are not key IKK targets, implicating other as yet undefined IKK targets to play a role in this process.
Copyright © 2022 The Authors.
Figures





Similar articles
-
A balance between B cell receptor and inhibitory receptor signaling controls plasma cell differentiation by maintaining optimal Ets1 levels.J Immunol. 2014 Jul 15;193(2):909-920. doi: 10.4049/jimmunol.1400666. Epub 2014 Jun 13. J Immunol. 2014. PMID: 24929000 Free PMC article.
-
Genetic Interaction between Lyn, Ets1, and Btk in the Control of Antibody Levels.J Immunol. 2015 Sep 1;195(5):1955-63. doi: 10.4049/jimmunol.1500165. Epub 2015 Jul 24. J Immunol. 2015. PMID: 26209625 Free PMC article.
-
Critical role of class IA PI3K for c-Rel expression in B lymphocytes.Blood. 2009 Jan 29;113(5):1037-44. doi: 10.1182/blood-2008-06-163725. Epub 2008 Oct 16. Blood. 2009. PMID: 18927440
-
Regulation of B-cell activation and differentiation by the phosphatidylinositol 3-kinase and phospholipase Cgamma pathway.Immunol Rev. 2000 Aug;176:30-46. doi: 10.1034/j.1600-065x.2000.00611.x. Immunol Rev. 2000. PMID: 11043766 Review.
-
Regulation and function of IKK and IKK-related kinases.Sci STKE. 2006 Oct 17;2006(357):re13. doi: 10.1126/stke.3572006re13. Sci STKE. 2006. PMID: 17047224 Review.
Cited by
-
Sequences within and upstream of the mouse Ets1 gene drive high level expression in B cells, but are not sufficient for consistent expression in T cells.bioRxiv [Preprint]. 2024 Aug 5:2024.08.02.606433. doi: 10.1101/2024.08.02.606433. bioRxiv. 2024. Update in: PLoS One. 2025 Mar 07;20(3):e0308896. doi: 10.1371/journal.pone.0308896. PMID: 39149372 Free PMC article. Updated. Preprint.
-
Sequences within and upstream of the mouse Ets1 gene drive high level expression in B cells, but are not sufficient for consistent expression in T cells.PLoS One. 2025 Mar 7;20(3):e0308896. doi: 10.1371/journal.pone.0308896. eCollection 2025. PLoS One. 2025. PMID: 40053568 Free PMC article.
References
-
- Nguyen HV, Mouly E, Chemin K, Luinaud R, Despres R, Fermand JP, Arnulf B, and Bories JC. 2012. The Ets-1 transcription factor is required for Stat1-mediated T-bet expression and IgG2a class switching in mouse B cells. Blood 119: 4174–4181. - PubMed
-
- Wang D, John SA, Clements JL, Percy DH, Barton KP, and Garrett-Sinha LA. 2005. Ets-1 deficiency leads to altered B cell differentiation, hyperresponsiveness to TLR9 and autoimmune disease. Int Immunol 17: 1179–1191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
