

Arsenic trioxide elicits prophylactic and therapeutic immune responses against solid tumors by inducing necroptosis and ferroptosis
- PMID: 36447031
- PMCID: PMC9794749
- DOI: 10.1038/s41423-022-00956-0
Arsenic trioxide elicits prophylactic and therapeutic immune responses against solid tumors by inducing necroptosis and ferroptosis
Abstract
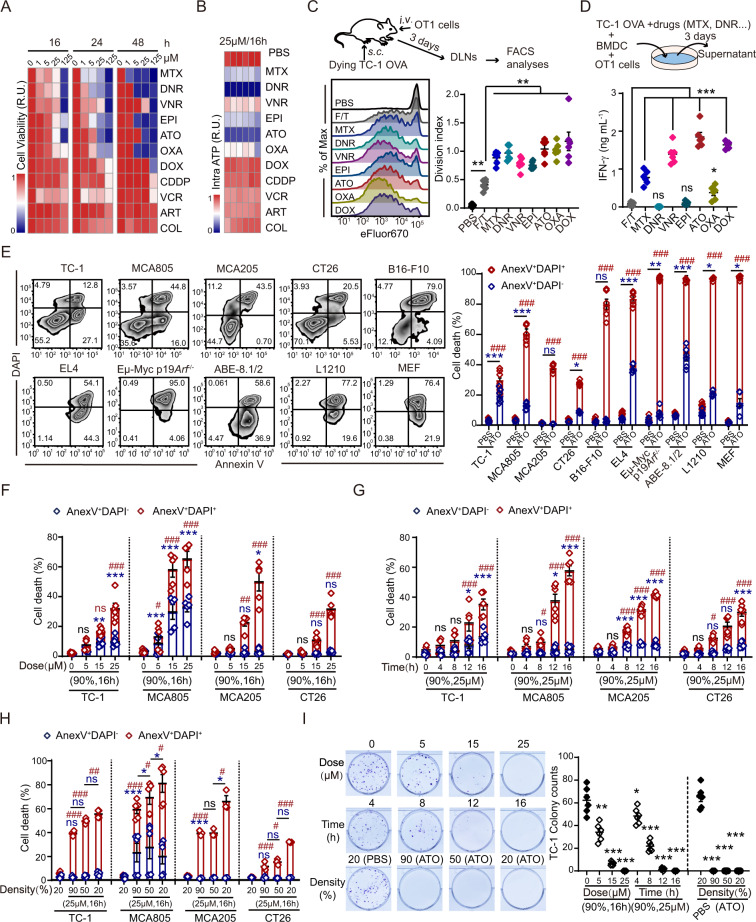
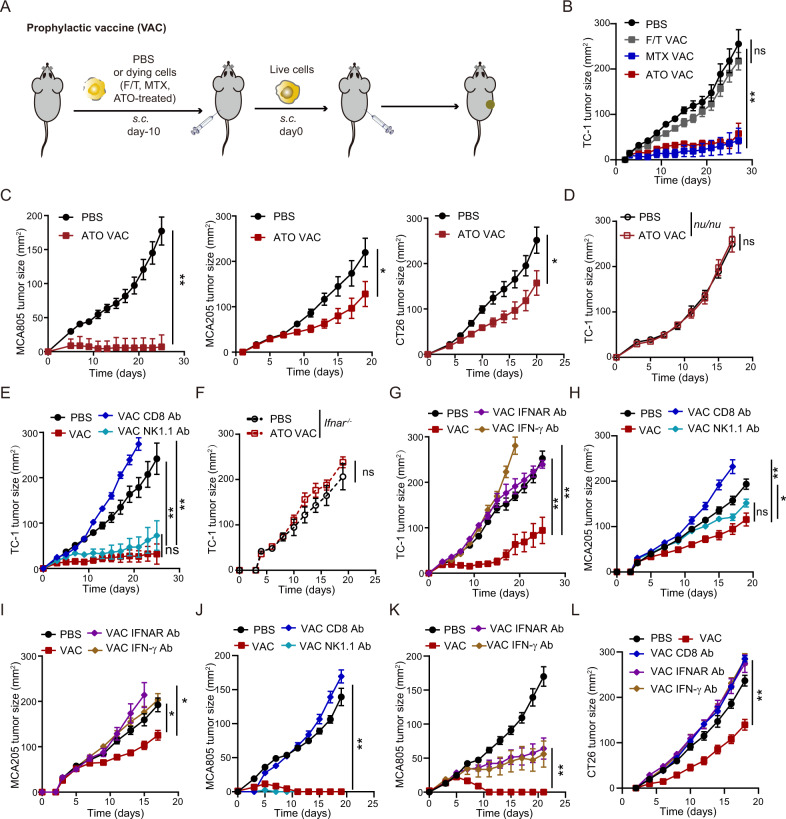
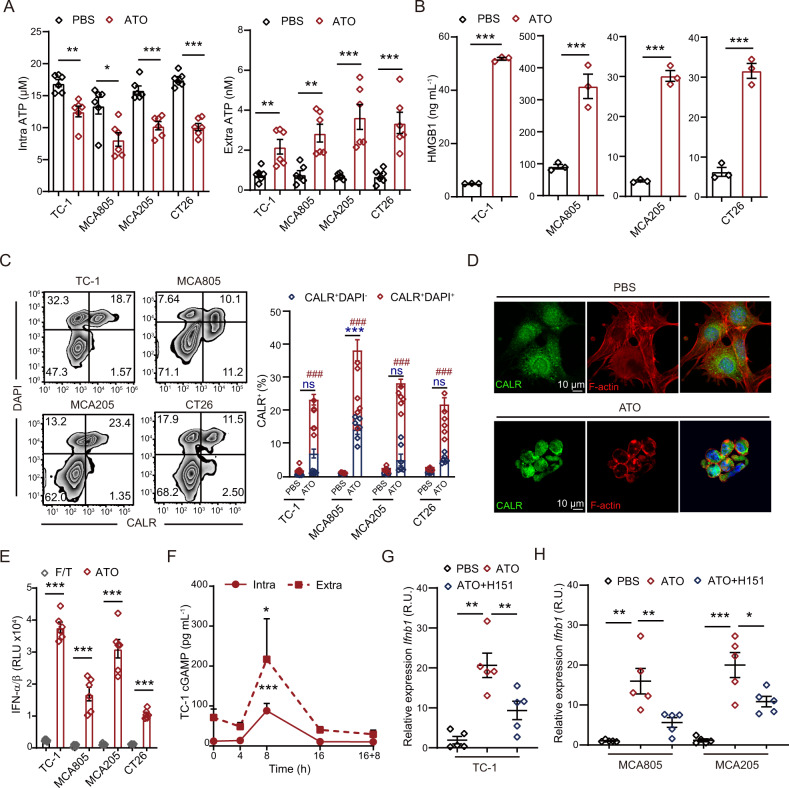
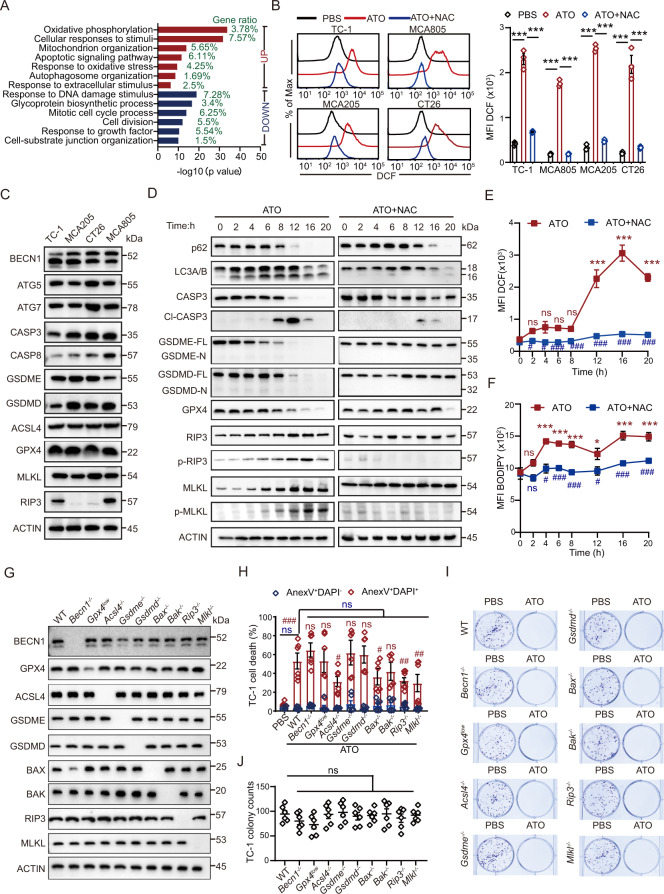
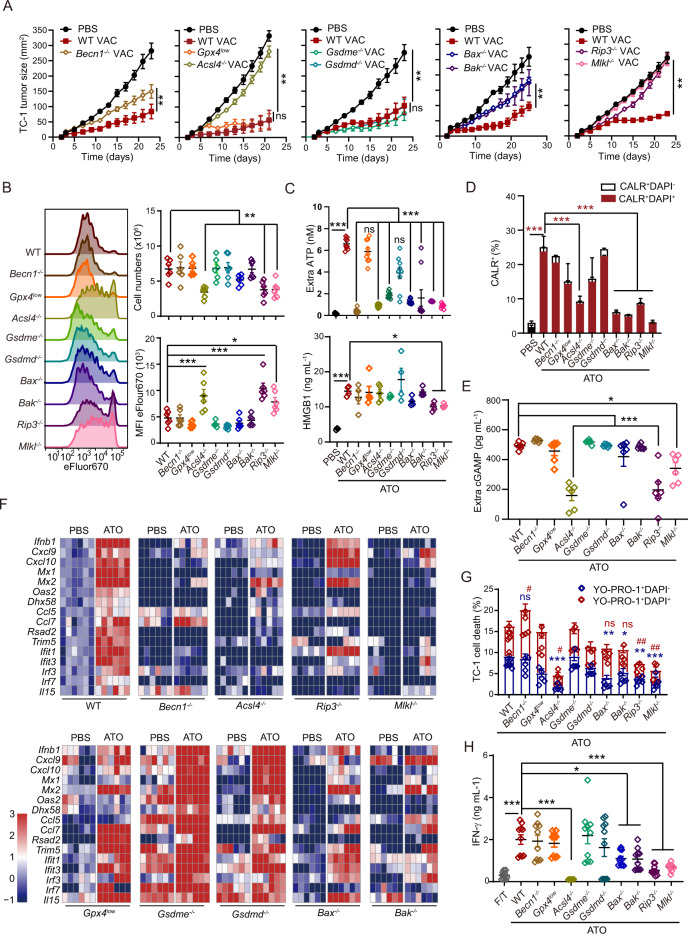
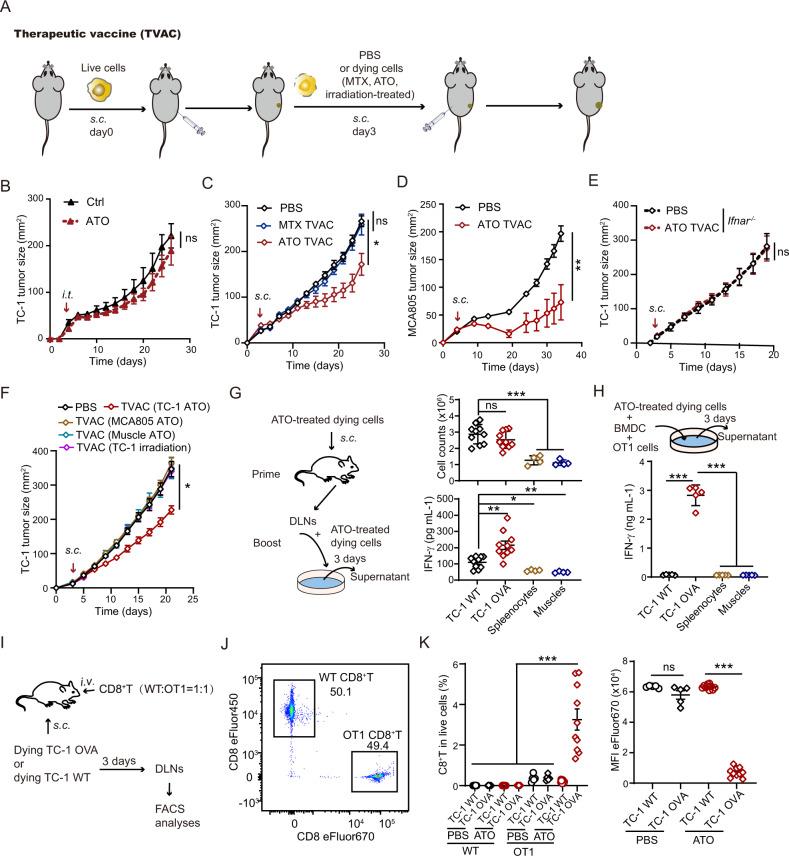
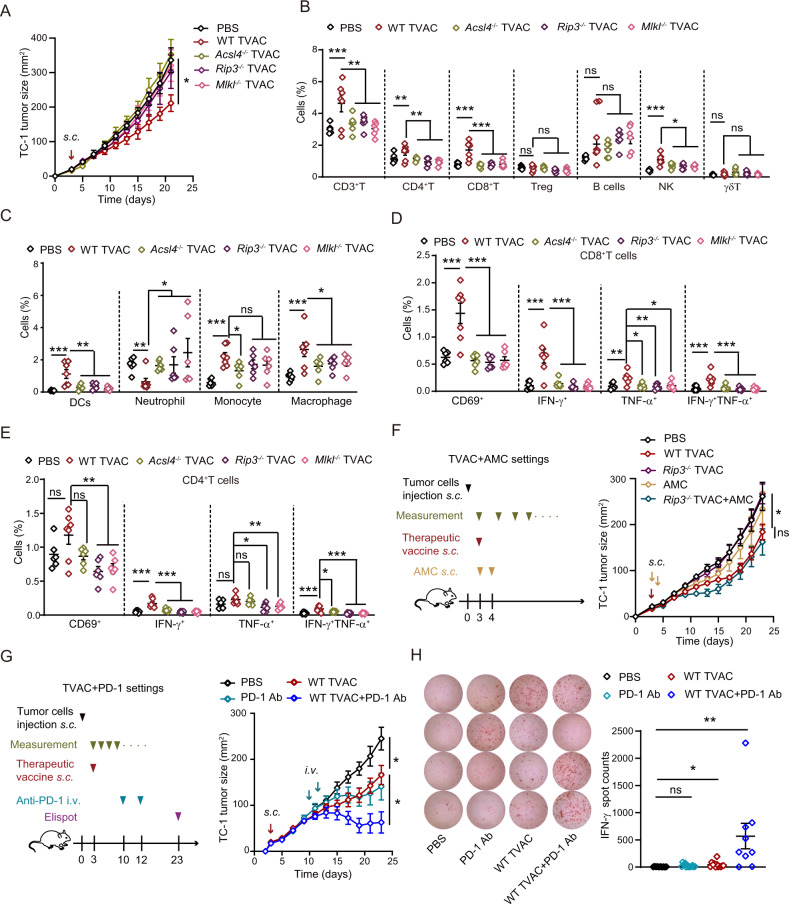
Boosting tumor immunosurveillance with vaccines has been proven to be a feasible and cost-effective strategy to fight cancer. Although major breakthroughs have been achieved in preventative tumor vaccines targeting oncogenic viruses, limited advances have been made in curative vaccines for virus-irrelevant malignancies. Accumulating evidence suggests that preconditioning tumor cells with certain cytotoxic drugs can generate whole-cell tumor vaccines with strong prophylactic activities. However, the immunogenicity of these vaccines is not sufficient to restrain the outgrowth of existing tumors. In this study, we identified arsenic trioxide (ATO) as a wide-spectrum cytotoxic and highly immunogenic drug through multiparameter screening. ATO preconditioning could generate whole-cell tumor vaccines with potent antineoplastic effects in both prophylactic and therapeutic settings. The tumor-preventive or tumor-suppressive benefits of these vaccines relied on CD8+ T cells and type I and II interferon signaling and could be linked to the release of immunostimulatory danger molecules. Unexpectedly, following ATO-induced oxidative stress, multiple cell death pathways were activated, including autophagy, apoptosis, necroptosis, and ferroptosis. CRISPR‒Cas9-mediated knockout of cell death executors revealed that the absence of Rip3, Mlkl, or Acsl4 largely abolished the efficacy of ATO-based prophylactic and therapeutic cancer vaccines. This therapeutic failure could be rescued by coadministration of danger molecule analogs. In addition, PD-1 blockade synergistically improved the therapeutic efficacy of ATO-based cancer vaccines by augmenting local IFN-γ production.
Keywords: Arsenic trioxide; Ferroptosis; Immunosurveillance; Necroptosis; Tumor vaccine.
© 2022. The Author(s), under exclusive licence to CSI and USTC.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Immunological consequences of arsenic trioxide-induced necrosis.Cell Mol Immunol. 2023 Mar;20(3):308-309. doi: 10.1038/s41423-023-00976-4. Epub 2023 Jan 25. Cell Mol Immunol. 2023. PMID: 36693921 Free PMC article. No abstract available.
References
-
- Finn OJ. The dawn of vaccines for cancer prevention. Nat Rev Immunol. 2018;18:183–94. 10.1038/nri.2017.140 - PubMed
-
- McMahon BJ, Bulkow LR, Singleton RJ, Williams J, Snowball M, Homan C, et al. Elimination of hepatocellular carcinoma and acute hepatitis B in children 25 years after a hepatitis B newborn and catch-up immunization program. Hepatology. 2011;54:801–7. 10.1002/hep.24442 - PubMed
-
- Sun X, Zeng L, Huang Y. Transcutaneous delivery of DNA/mRNA for cancer therapeutic vaccination. J Gene Med. 2019;21:e3089. 10.1002/jgm.3089 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
