

Bridged Proteolysis Targeting Chimera (PROTAC) Enables Degradation of Undruggable Targets
- PMID: 36448571
- PMCID: PMC9772293
- DOI: 10.1021/jacs.2c09255
Bridged Proteolysis Targeting Chimera (PROTAC) Enables Degradation of Undruggable Targets
Abstract
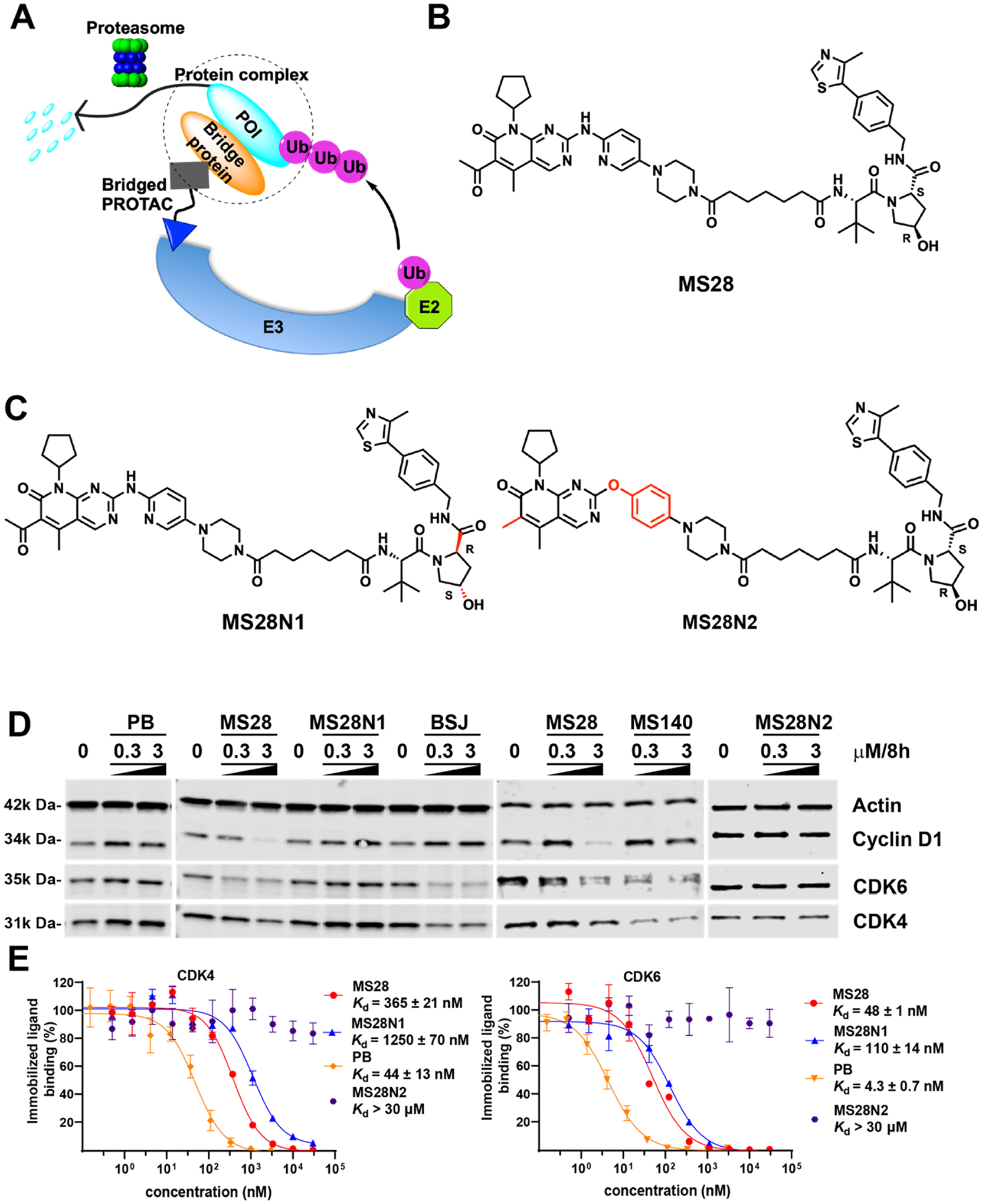
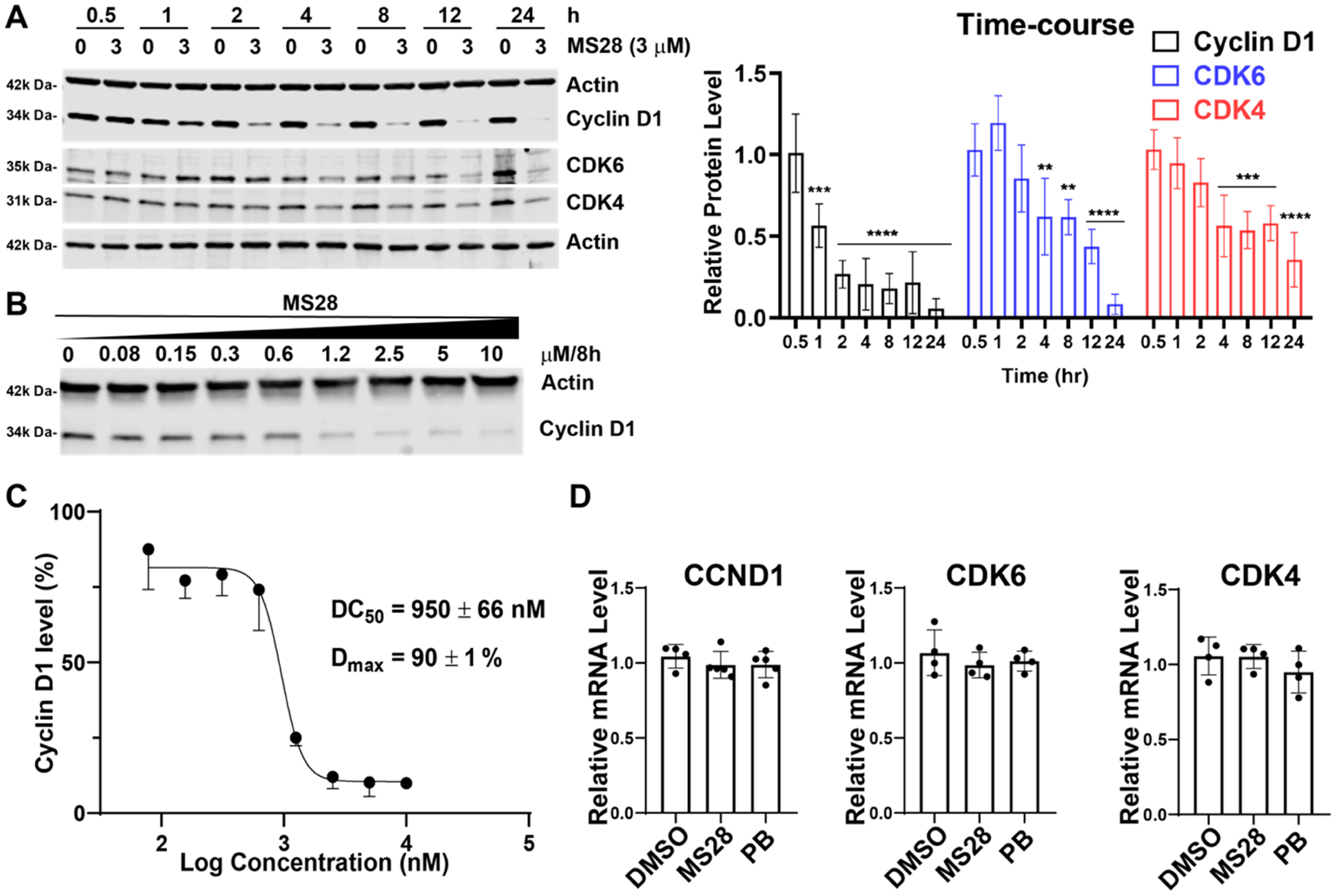
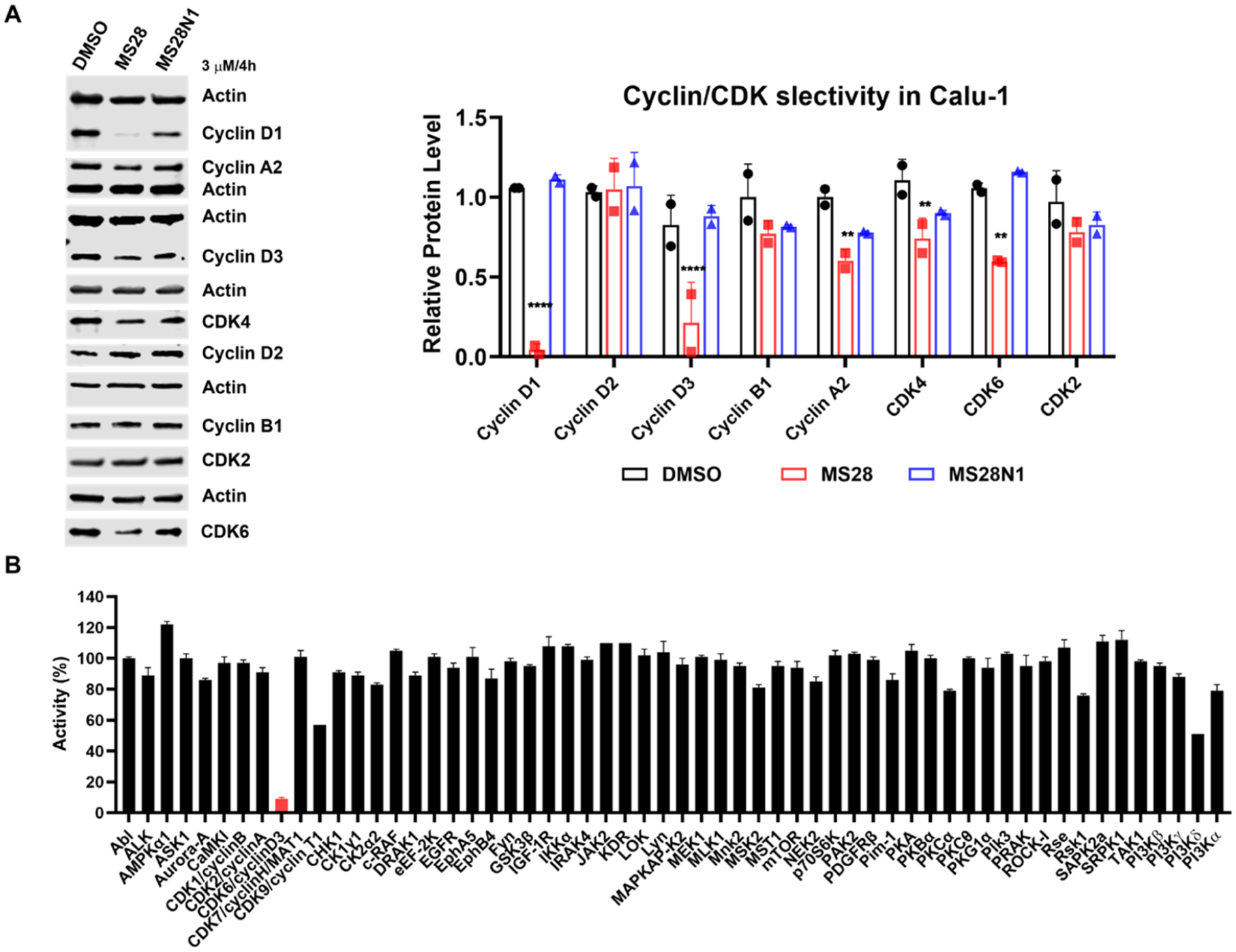
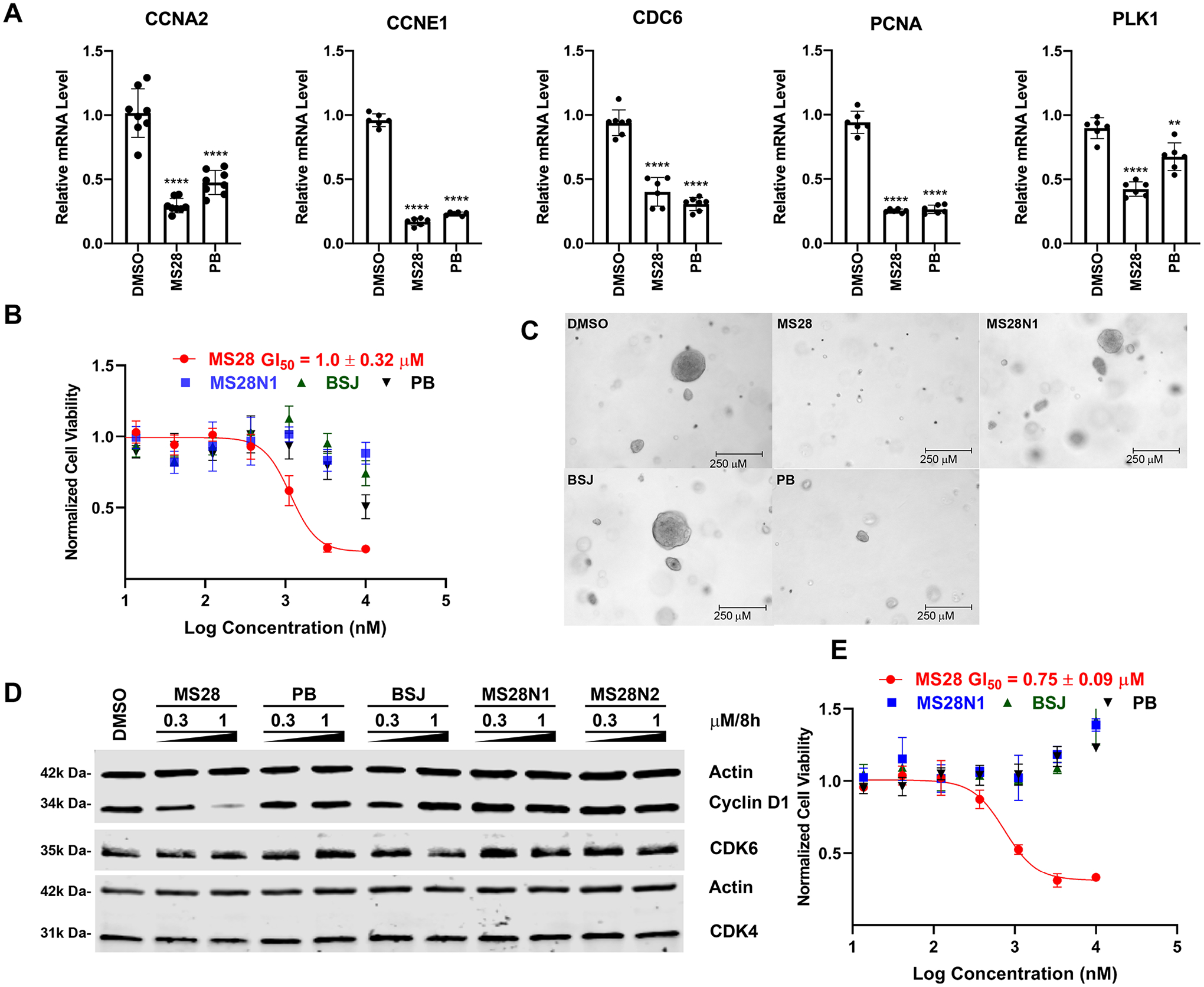
Proteolysis Targeting Chimeras (PROTACs) are attractive therapeutic modalities for degrading disease-causing proteins. While many PROTACs have been developed for numerous protein targets, current small-molecule PROTAC approaches cannot target undruggable proteins that do not have small-molecule binders. Here, we present a novel PROTAC approach, termed bridged PROTAC, which utilizes a small-molecule binder of the target protein's binding partner to recruit the protein complex into close proximity with an E3 ubiquitin ligase to target undruggable proteins. Applying this bridged PROTAC strategy, we discovered MS28, the first-in-class degrader of cyclin D1, which lacks a small-molecule binder. MS28 effectively degrades cyclin D1, with faster degradation kinetics and superior degradation efficiency than CDK4/6, through recruiting the CDK4/6-cyclin D1 complex to the von Hippel-Lindau E3 ligase. MS28 also suppressed the proliferation of cancer cells more effectively than CDK4/6 inhibitors and degraders. Altogether, the bridged PROTAC strategy could provide a generalizable platform for targeting undruggable proteins.
Conflict of interest statement
The authors declare the following competing financial interest(s): The Jin laboratory received research funds from Celgene Corporation, Levo Therapeutics, Inc., Cullgen, Inc. and Cullinan Oncology, Inc. J.J. is a cofounder and equity shareholder in Cullgen, Inc. and a consultant for Cullgen, Inc., EpiCypher, Inc., and Accent Therapeutics, Inc.
Figures

Similar articles
-
Discovery and Characterization of a Novel Cereblon-Recruiting PRC1 Bridged PROTAC Degrader.J Med Chem. 2024 Apr 25;67(8):6880-6892. doi: 10.1021/acs.jmedchem.4c00538. Epub 2024 Apr 12. J Med Chem. 2024. PMID: 38607318 Free PMC article.
-
Journey of Von Hippel-Lindau (VHL) E3 ligase in PROTACs design: From VHL ligands to VHL-based degraders.Eur J Med Chem. 2024 Feb 5;265:116041. doi: 10.1016/j.ejmech.2023.116041. Epub 2023 Dec 14. Eur J Med Chem. 2024. PMID: 38199162 Review.
-
Harnessing the SPOP E3 Ubiquitin Ligase via a Bridged Proteolysis Targeting Chimera (PROTAC) Strategy for Targeted Protein Degradation.J Med Chem. 2025 Apr 24;68(8):8634-8647. doi: 10.1021/acs.jmedchem.5c00295. Epub 2025 Apr 9. J Med Chem. 2025. PMID: 40202531
-
Leveraging Dual-Ligase Recruitment to Enhance Protein Degradation via a Heterotrivalent Proteolysis Targeting Chimera.J Am Chem Soc. 2024 Dec 11;146(49):33675-33711. doi: 10.1021/jacs.4c11556. Epub 2024 Nov 28. J Am Chem Soc. 2024. PMID: 39606859 Free PMC article.
-
[Advances in the preclinical and clinical research of proteolysis targeting chimera].Sheng Wu Gong Cheng Xue Bao. 2023 Sep 25;39(9):3615-3627. doi: 10.13345/j.cjb.221040. Sheng Wu Gong Cheng Xue Bao. 2023. PMID: 37805842 Review. Chinese.
Cited by
-
Coming of Age: Targeting Cyclin K in Cancers.Cells. 2023 Aug 11;12(16):2044. doi: 10.3390/cells12162044. Cells. 2023. PMID: 37626854 Free PMC article. Review.
-
Targeting N-Myc in neuroblastoma with selective Aurora kinase A degraders.Cell Chem Biol. 2025 Feb 20;32(2):352-362.e10. doi: 10.1016/j.chembiol.2024.12.006. Epub 2025 Jan 7. Cell Chem Biol. 2025. PMID: 39778578
-
p53 in colorectal cancer: from a master player to a privileged therapy target.J Transl Med. 2025 Jun 19;23(1):684. doi: 10.1186/s12967-025-06566-4. J Transl Med. 2025. PMID: 40537809 Free PMC article. Review.
-
Non-coding RNA and drug resistance in head and neck cancer.Cancer Drug Resist. 2024 Sep 20;7:34. doi: 10.20517/cdr.2024.59. eCollection 2024. Cancer Drug Resist. 2024. PMID: 39403599 Free PMC article. Review.
-
Targeting CDKs in cancer therapy: advances in PROTACs and molecular glues.NPJ Precis Oncol. 2025 Jun 28;9(1):204. doi: 10.1038/s41698-025-00931-8. NPJ Precis Oncol. 2025. PMID: 40581698 Free PMC article. Review.
References
-
- Schapira M; Calabrese MF; Bullock AN; Crews CM Targeted protein degradation: expanding the toolbox. Nat. Rev. Drug Discovery 2019, 18, 949–963. - PubMed
-
- Bai L; Zhou H; Xu R; Zhao Y; Chinnaswamy K; McEachern D; Chen J; Yang C-Y; Liu Z; Wang M; Liu L; Jiang H; Wen B; Kumar P; Meagher JL; Sun D; Stuckey JA; Wang S A potent and selective small-molecule degrader of STAT3 achieves complete tumor regression in vivo. Cancer Cell 2019, 36, 498–511.e17. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
