

Antigen-presenting T cells provide critical B7 co-stimulation for thymic iNKT cell development via CD28-dependent trogocytosis
- PMID: 36450247
- PMCID: PMC9805342
- DOI: 10.1016/j.celrep.2022.111731
Antigen-presenting T cells provide critical B7 co-stimulation for thymic iNKT cell development via CD28-dependent trogocytosis
Abstract
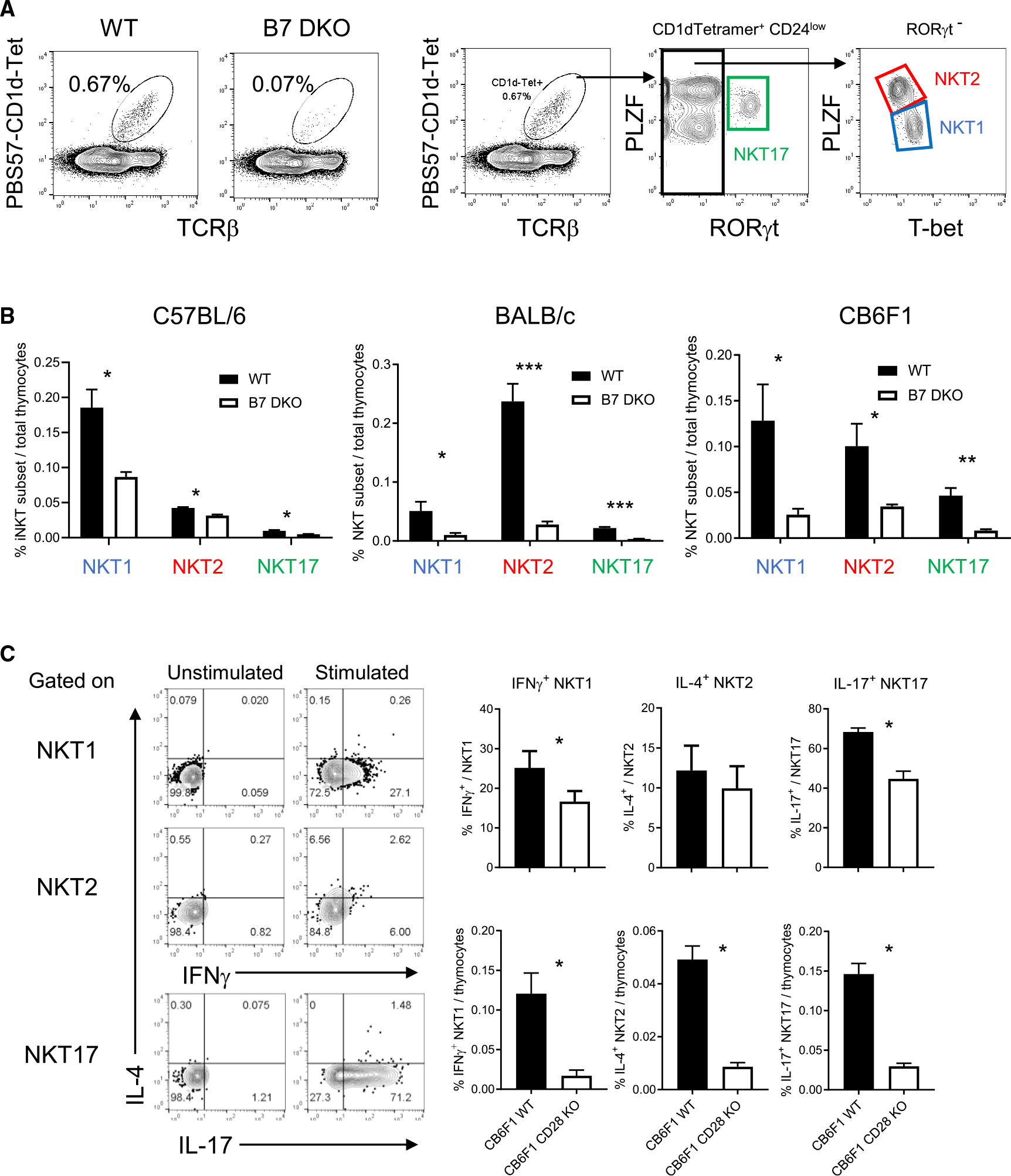
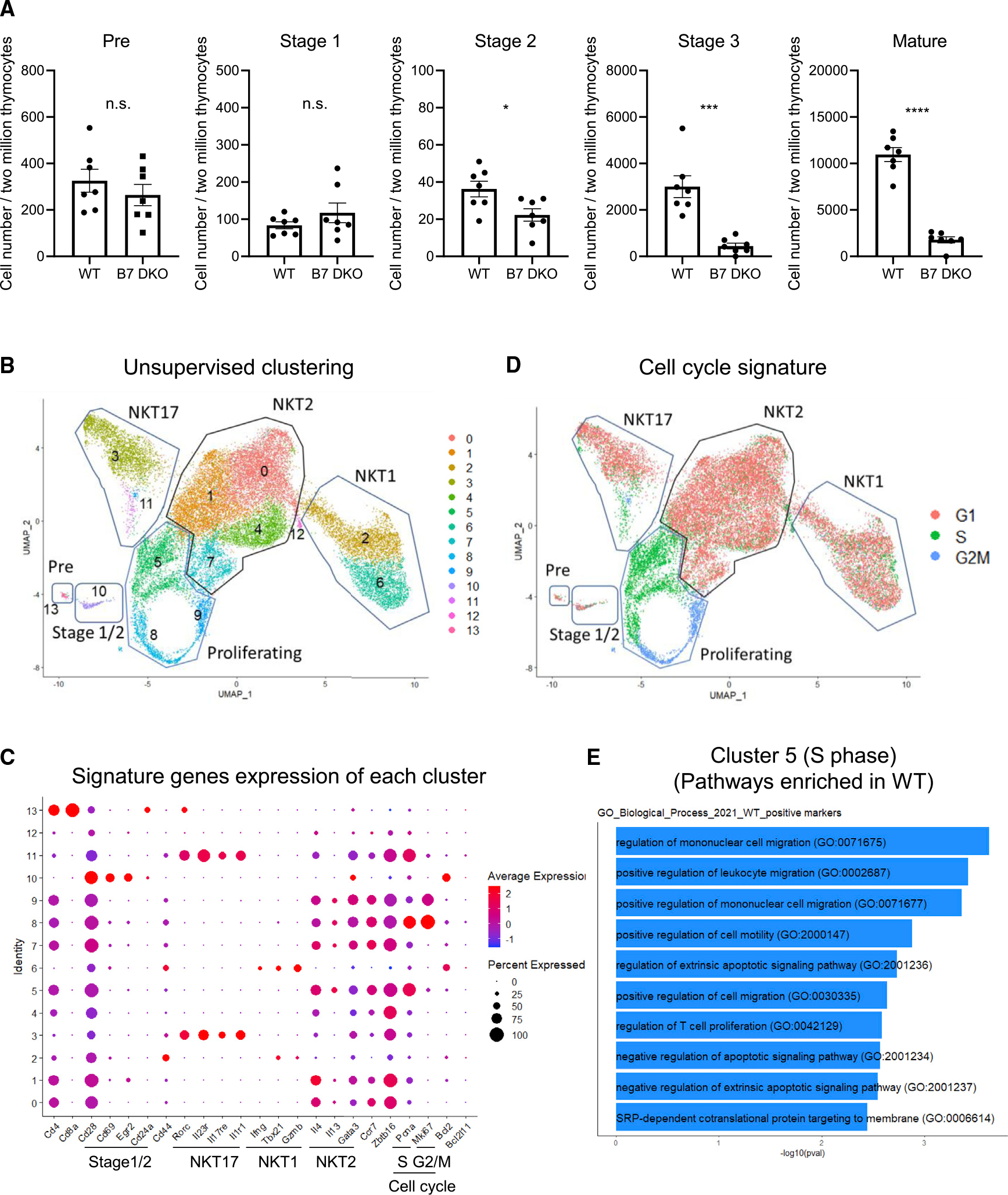
Invariant natural killer T (iNKT) cell development in the thymus depends on T cell receptor recognition of CD1d ligand on CD4/CD8 double-positive thymocytes. We previously reported that B7-CD28 co-stimulation is required for thymic iNKT cell development, but the cellular and molecular mechanisms underlying this co-stimulatory requirement are not understood. Here we report that CD28 expression on CD1d-expressing antigen-presenting T cells is required for thymic iNKT cell development. Mechanistically, antigen-presenting T cells provide co-stimulation through an unconventional mechanism, acquiring B7 molecules via CD28-dependent trogocytosis from B7-expressing thymic epithelial cells, dendritic cells, and B cells and providing critical B7 co-stimulation to developing iNKT cells. Thus, the present study demonstrates a mechanism of B7 co-stimulation in thymic T cell development by antigen-presenting T cells.
Keywords: B7-CD28 co-stimulation; CP: Immunology; antigen-presenting T cell; iNKT; thymic T cell development; trogocytosis.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
