

Redirecting tropane alkaloid metabolism reveals pyrrolidine alkaloid diversity in Atropa belladonna
- PMID: 36451537
- PMCID: PMC10107824
- DOI: 10.1111/nph.18651
Redirecting tropane alkaloid metabolism reveals pyrrolidine alkaloid diversity in Atropa belladonna
Abstract
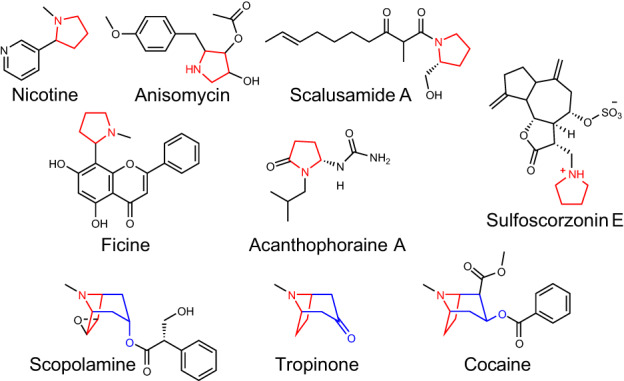
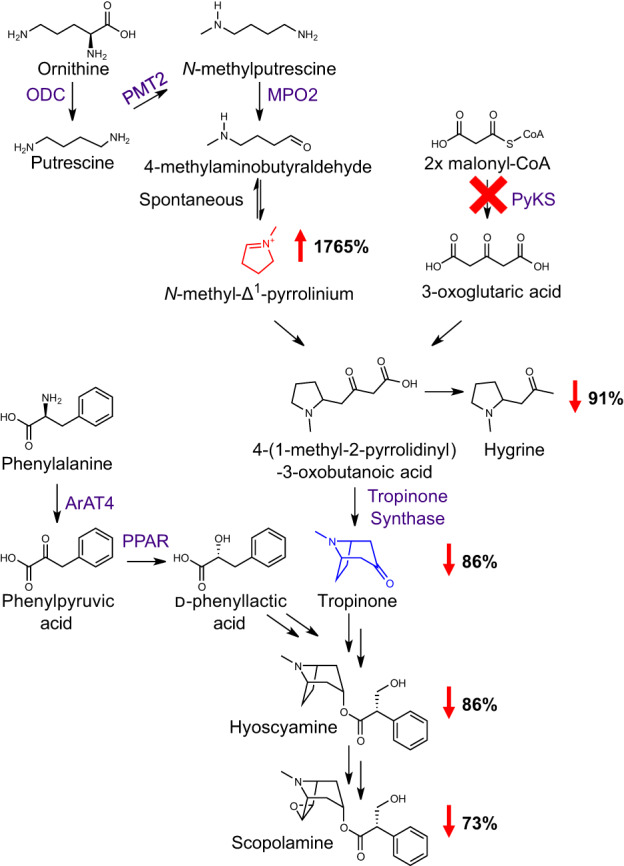
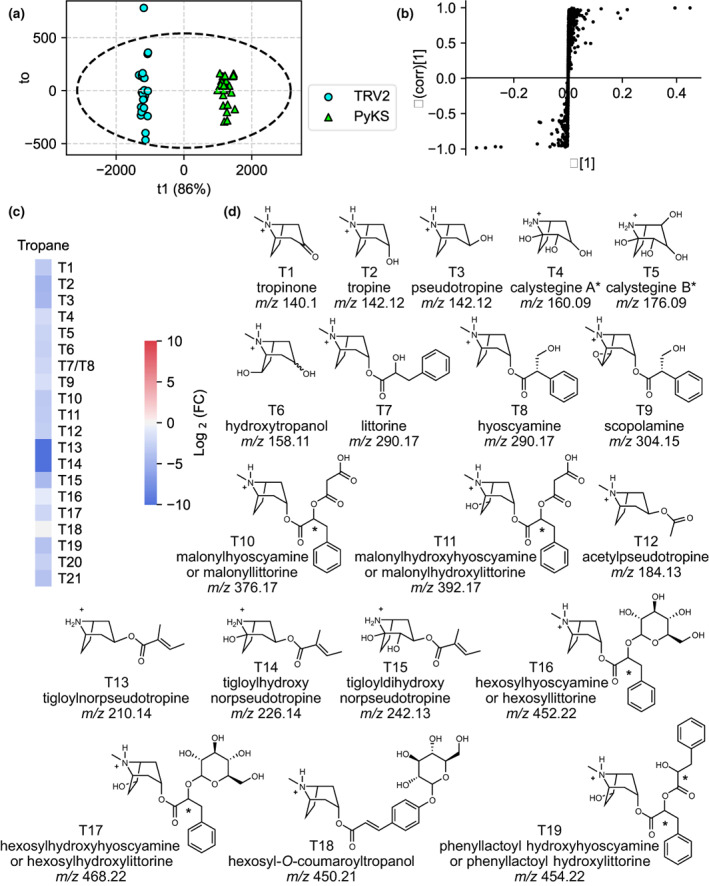
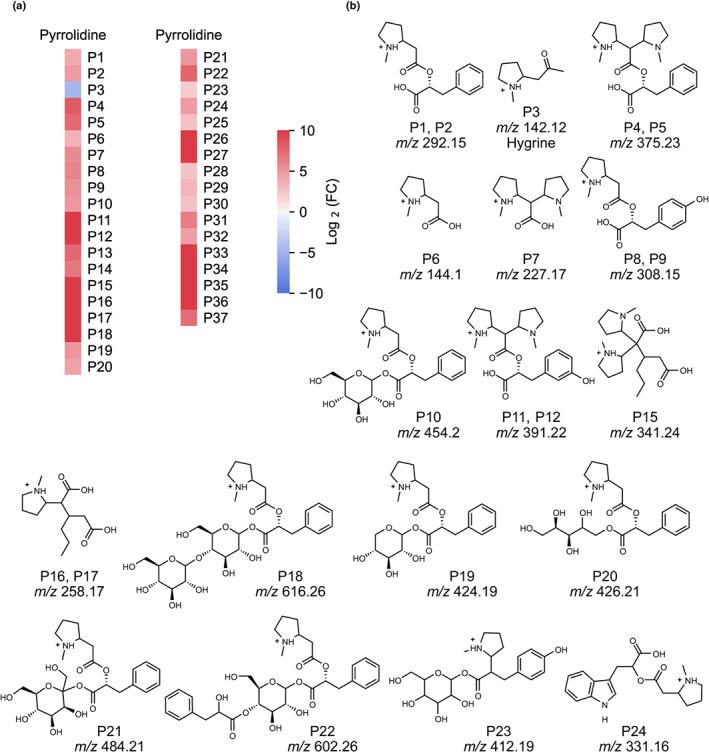
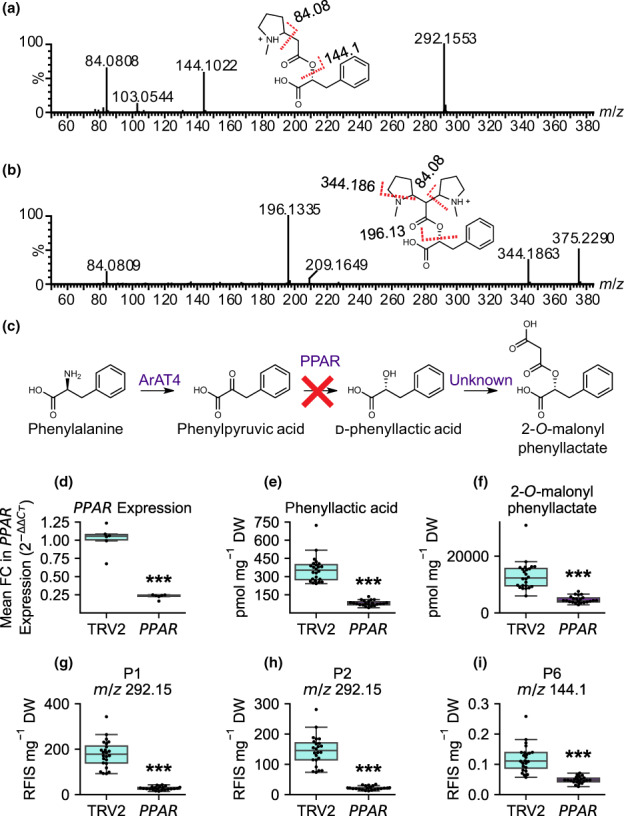
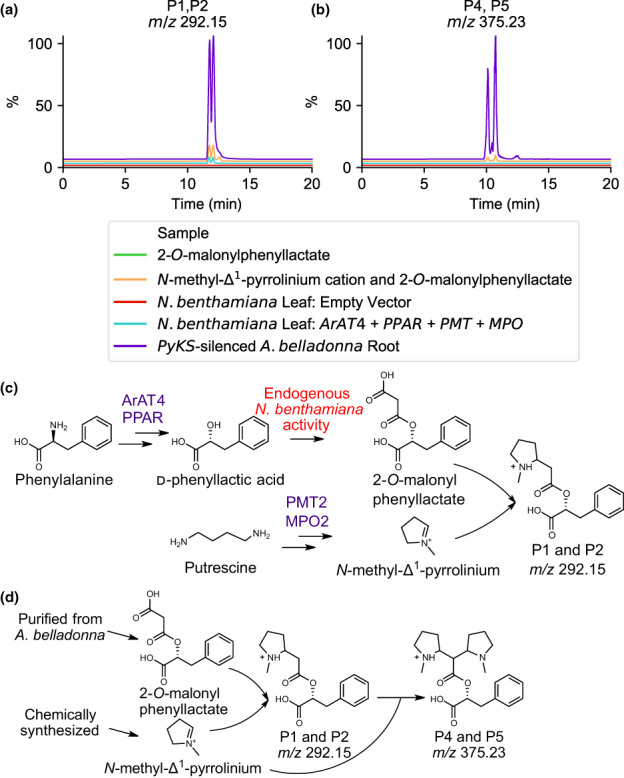
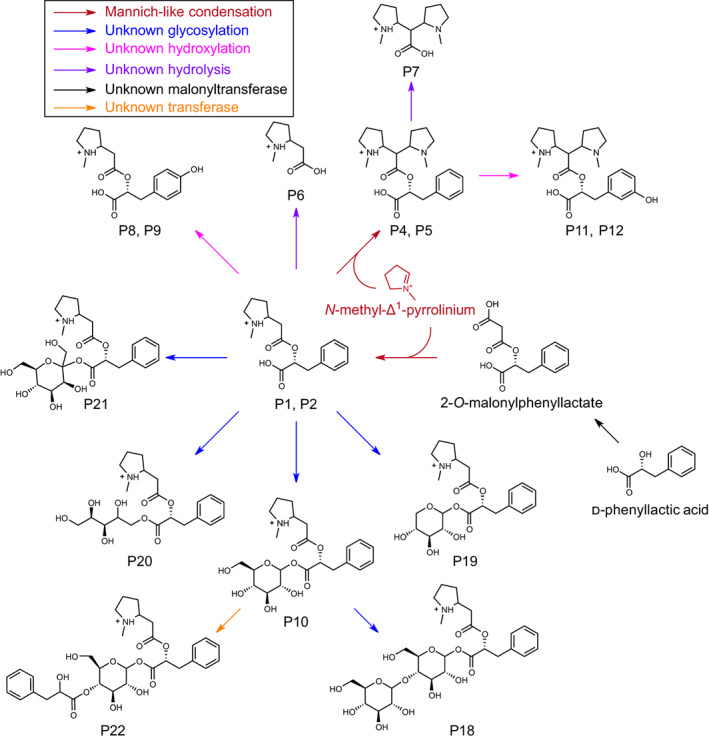
Plant-specialized metabolism is complex, with frequent examples of highly branched biosynthetic pathways, and shared chemical intermediates. As such, many plant-specialized metabolic networks are poorly characterized. The N-methyl Δ1 -pyrrolinium cation is a simple pyrrolidine alkaloid and precursor of pharmacologically important tropane alkaloids. Silencing of pyrrolidine ketide synthase (AbPyKS) in the roots of Atropa belladonna (Deadly Nightshade) reduces tropane alkaloid abundance and causes high N-methyl Δ1 -pyrrolinium cation accumulation. The consequences of this metabolic shift on alkaloid metabolism are unknown. In this study, we utilized discovery metabolomics coupled with AbPyKS silencing to reveal major changes in the root alkaloid metabolome of A. belladonna. We discovered and annotated almost 40 pyrrolidine alkaloids that increase when AbPyKS activity is reduced. Suppression of phenyllactate biosynthesis, combined with metabolic engineering in planta, and chemical synthesis indicates several of these pyrrolidines share a core structure formed through the nonenzymatic Mannich-like decarboxylative condensation of the N-methyl Δ1 -pyrrolinium cation with 2-O-malonylphenyllactate. Decoration of this core scaffold through hydroxylation and glycosylation leads to mono- and dipyrrolidine alkaloid diversity. This study reveals the previously unknown complexity of the A. belladonna root metabolome and creates a foundation for future investigation into the biosynthesis, function, and potential utility of these novel alkaloids.
Keywords: Atropa belladonna; Solanaceae; metabolomics; nonenzymatic catalysis; pyrrolidine alkaloids; specialized metabolism; tropane alkaloids.
© 2022 The Authors New Phytologist © 2022 New Phytologist Foundation.
Conflict of interest statement
None declared.
Figures
References
-
- Bedewitz MA, Gongora‐Castillo E, Uebler JB, Gonzales‐Vigil E, Wiegert‐Rininger KE, Childs KL, Hamilton JP, Vaillancourt B, Yeo YS, Chappell J et al. 2014. A root‐expressed l‐phenylalanine:4‐hydroxyphenylpyruvate aminotransferase is required for tropane alkaloid biosynthesis in Atropa belladonna . Plant Cell 26: 3745–3762. - PMC - PubMed
-
- Blair LM, Calvert MB, Sperry J. 2017. Flavoalkaloids‐isolation, biological activity, and total synthesis. The Alkaloids: Chemistry and Biology 77: 85–115. - PubMed
-
- Dieuleveux V, Lemarinier S, Gueguen M. 1998. Antimicrobial spectrum and target site of D‐3‐phenyllactic acid. International Journal of Food Microbiology 40: 177–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
