

SOX4 promotes beige adipocyte-mediated adaptive thermogenesis by facilitating PRDM16-PPARγ complex
- PMID: 36451857
- PMCID: PMC9706582
- DOI: 10.7150/thno.77102
SOX4 promotes beige adipocyte-mediated adaptive thermogenesis by facilitating PRDM16-PPARγ complex
Abstract
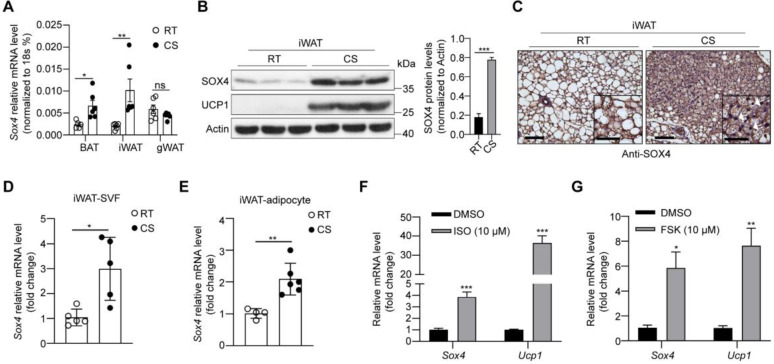
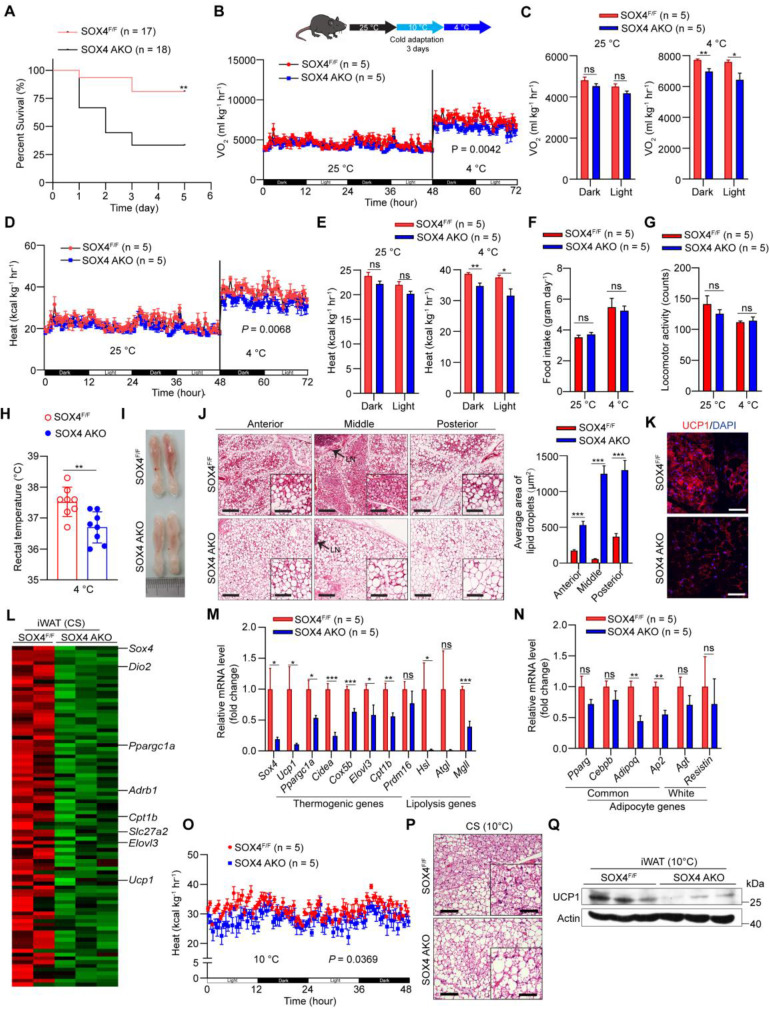
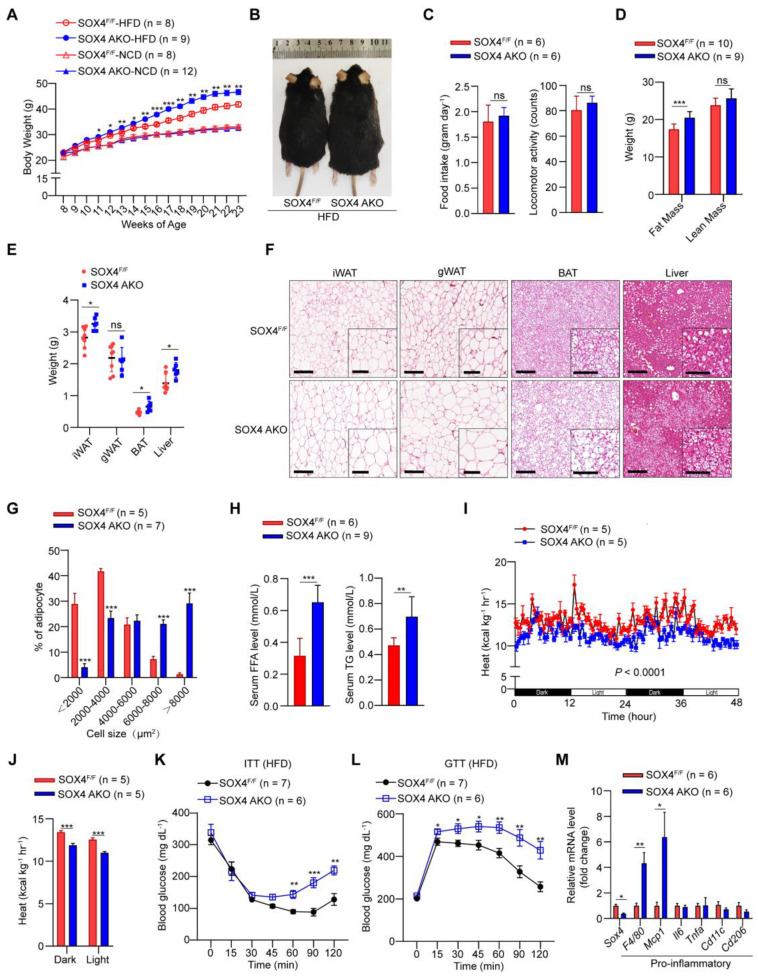
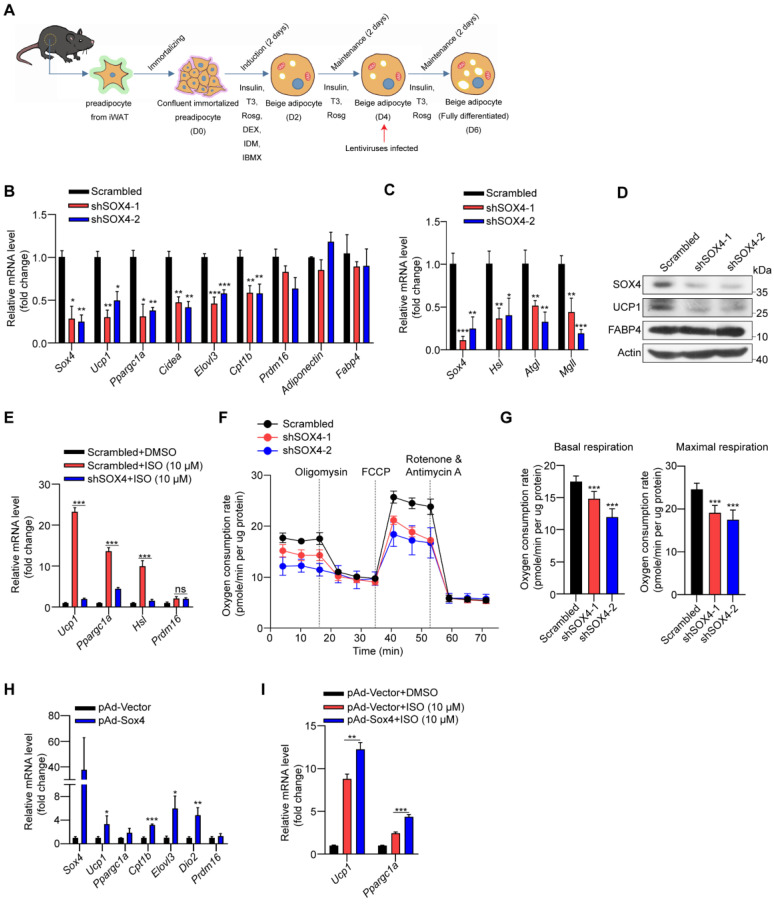
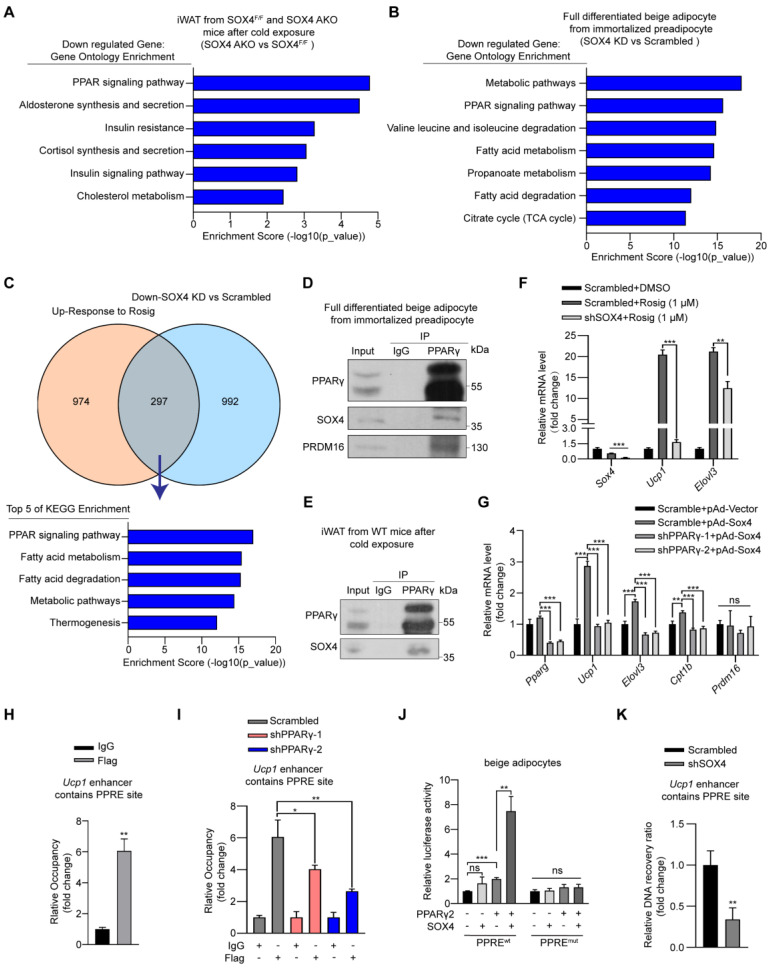
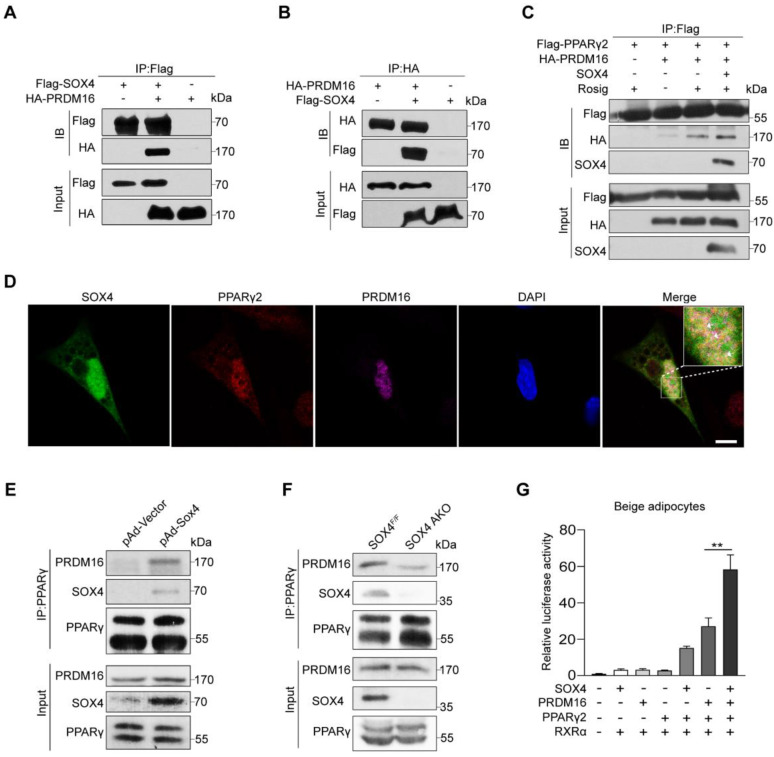
Brown and beige fat protect against cold environments and obesity by catabolizing stored energy to generate heat. This process is achieved by controlling thermogenesis-related gene expression and the development of brown/beige fat through the induction of transcription factors, most notably PPARγ. However, the cofactors that induce the expression of thermogenic genes with PPARγ are still not well understood. In this study, we explored the role of SOX4 in adaptive thermogenesis and its relationship with PPARγ. Methods: Whole transcriptome deep sequencing (RNA-seq) analysis of inguinal subcutaneous white adipose tissue (iWAT) after cold stimulation was performed to identify genes with differential expression in mice. Indirect calorimetry detected oxygen consumption rate and heat generation. mRNA levels were analyzed by qPCR assays. Proteins were detected by immunoblotting and immunofluorescence. Interaction of proteins was detected by endogenous and exogenous Co-IP. ChIP-qPCR, FAIRE assay and luciferase reporter assays were used to investigate transcriptional regulation. Results: SOX4 was identified as the main transcriptional effector of thermogenesis. Mice with either adipocyte-specific or UCP1+ cells deletion of SOX4 exhibited significant cold intolerance, decreased energy expenditure, and beige adipocyte formation, which was attributed to decreased thermogenic gene expression. In addition, these mice developed obesity on a high-fat diet, with severe hepatic steatosis, insulin resistance, and inflammation. At the cell level, loss of SOX4 from preadipocytes inhibited the development of beige adipocytes, and loss of SOX4 from mature beige adipocytes reduced the expression of thermogenesis-related genes and energy metabolism. Mechanistically, SOX4 stimulated the transcriptional activity of Ucp1 by binding to PPARγ and activating its transcriptional function. These actions of SOX4 were, at least partly, mediated by recruiting PRDM16 to PPARγ, thus forming a transcriptional complex to elevate the expression of thermogenic genes. Conclusion: SOX4, as a coactivator of PPARγ, drives the thermogenic gene expression program and thermogenesis of beige fat, promoting energy expenditure. It has important physiological significance in resisting cold and obesity.
Keywords: PPARγ; SOX4.; beige fat; obesity; thermogenesis.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures
Similar articles
-
Cold acclimation and pioglitazone combined increase thermogenic capacity of brown and white adipose tissues but this does not translate into higher energy expenditure in mice.Am J Physiol Endocrinol Metab. 2023 Apr 1;324(4):E358-E373. doi: 10.1152/ajpendo.00217.2022. Epub 2023 Mar 1. Am J Physiol Endocrinol Metab. 2023. PMID: 36856189
-
Bone morphogenic protein 9 is a novel thermogenic hepatokine secreted in response to cold exposure.Metabolism. 2022 Apr;129:155139. doi: 10.1016/j.metabol.2022.155139. Epub 2022 Jan 19. Metabolism. 2022. PMID: 35063533
-
Id1 Promotes Obesity by Suppressing Brown Adipose Thermogenesis and White Adipose Browning.Diabetes. 2017 Jun;66(6):1611-1625. doi: 10.2337/db16-1079. Epub 2017 Mar 7. Diabetes. 2017. PMID: 28270523 Free PMC article.
-
Transcriptional Factors of Thermogenic Adipocyte Development and Generation of Brown and Beige Adipocytes From Stem Cells.Stem Cell Rev Rep. 2020 Oct;16(5):876-892. doi: 10.1007/s12015-020-10013-w. Stem Cell Rev Rep. 2020. PMID: 32728995 Review.
-
UCP1 Dependent and Independent Thermogenesis in Brown and Beige Adipocytes.Front Endocrinol (Lausanne). 2020 Jul 28;11:498. doi: 10.3389/fendo.2020.00498. eCollection 2020. Front Endocrinol (Lausanne). 2020. PMID: 32849287 Free PMC article. Review.
Cited by
-
Lactiplantibacillus plantarum N4 ameliorates lipid metabolism and gut microbiota structure in high fat diet-fed rats.Front Microbiol. 2024 Jun 7;15:1390293. doi: 10.3389/fmicb.2024.1390293. eCollection 2024. Front Microbiol. 2024. PMID: 38912346 Free PMC article.
-
Mechanisms of adipocyte regulation: Insights from HADHB gene modulation.PLoS One. 2025 Mar 27;20(3):e0319384. doi: 10.1371/journal.pone.0319384. eCollection 2025. PLoS One. 2025. PMID: 40146690 Free PMC article.
-
Galectin-1 Promotes Gastric Carcinoma Progression and Cisplatin Resistance Through the NRP-1/c-JUN/Wee1 Pathway.J Gastric Cancer. 2024 Jul;24(3):300-315. doi: 10.5230/jgc.2024.24.e25. J Gastric Cancer. 2024. PMID: 38960889 Free PMC article.
-
SOX4 facilitates brown fat development and maintenance through EBF2-mediated thermogenic gene program in mice.Cell Death Differ. 2025 Mar;32(3):447-465. doi: 10.1038/s41418-024-01397-0. Epub 2024 Oct 15. Cell Death Differ. 2025. PMID: 39402212 Free PMC article.
-
PRDM16 suppresses pyroptosis to attenuate the progression of AKI caused by rhabdomyolysis via upregulation of USP10.Cell Mol Life Sci. 2025 Apr 2;82(1):138. doi: 10.1007/s00018-025-05666-0. Cell Mol Life Sci. 2025. PMID: 40175611 Free PMC article.
References
-
- Chouchani ET, Kazak L, Spiegelman BM. New advances in adaptive thermogenesis: UCP1 and beyond. Cell Metab. 2019;29:27–37. - PubMed
-
- Harms M, Seale P. Brown and beige fat: development, function and therapeutic potential. Nat Med. 2013;19:1252–63. - PubMed
-
- Wang Y, Li X, Liu C, Zhou L, Shi L, Zhang Z, WTAP regulates postnatal development of brown adipose tissue by stabilizing METTL3 in mice. Life Metabolism. 2022. loac028.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
