

Grabody B, an IGF1 receptor-based shuttle, mediates efficient delivery of biologics across the blood-brain barrier
- PMID: 36452865
- PMCID: PMC9701613
- DOI: 10.1016/j.crmeth.2022.100338
Grabody B, an IGF1 receptor-based shuttle, mediates efficient delivery of biologics across the blood-brain barrier
Abstract
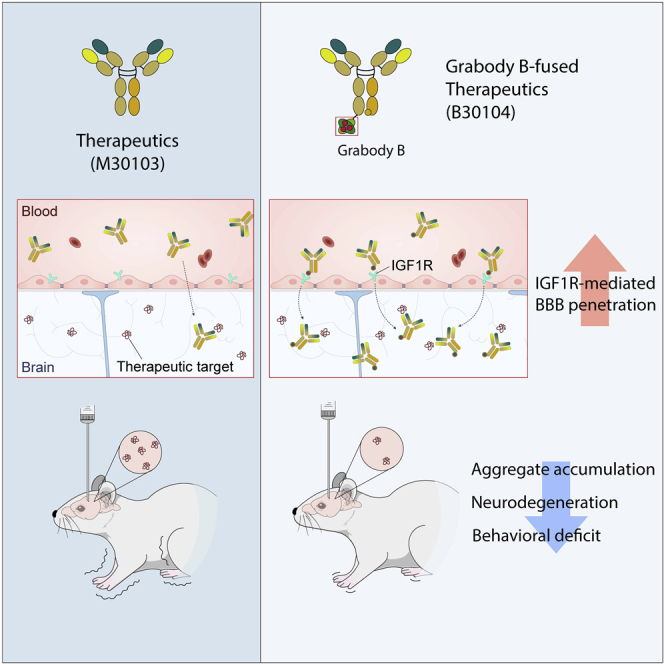
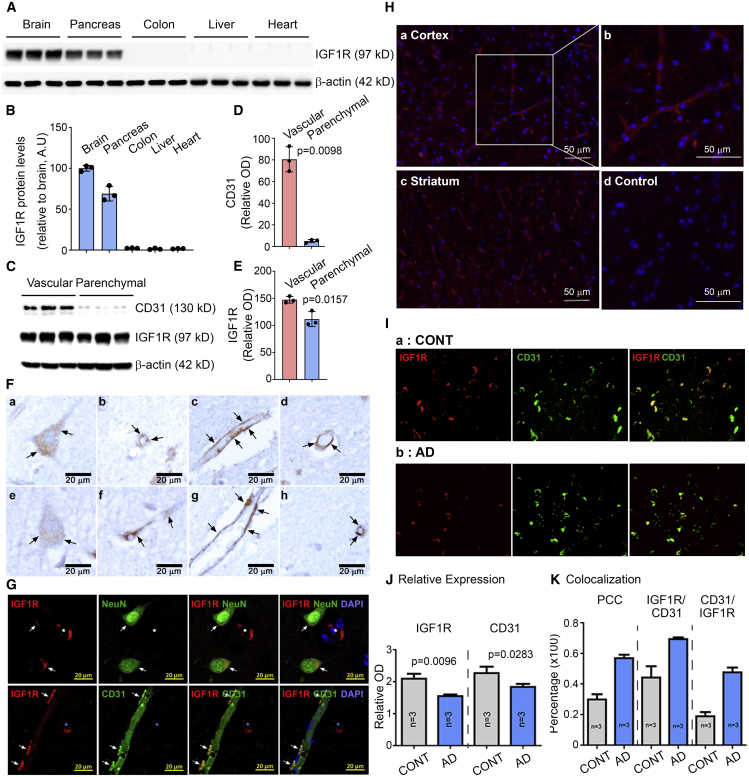
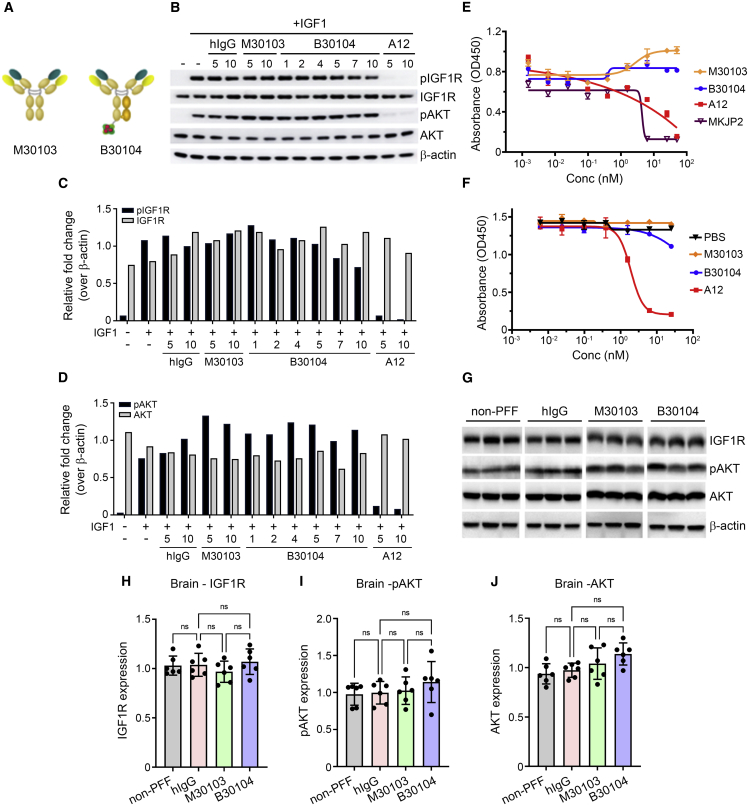
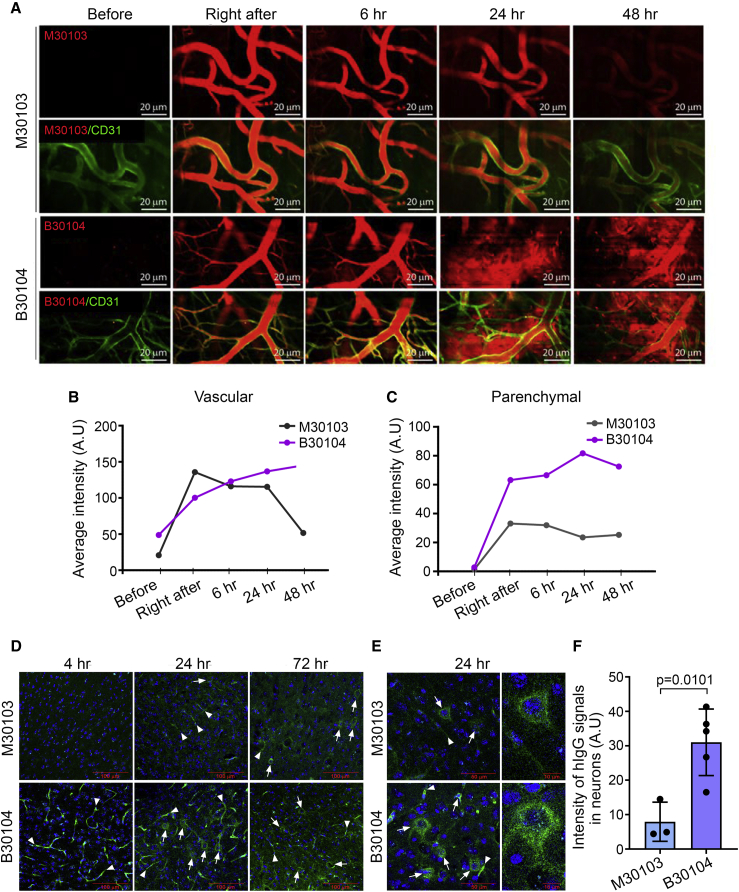
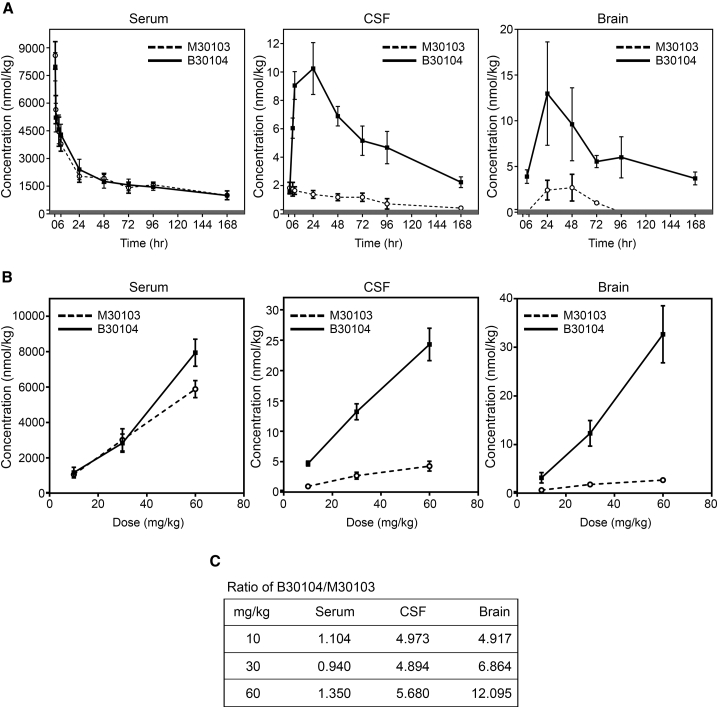
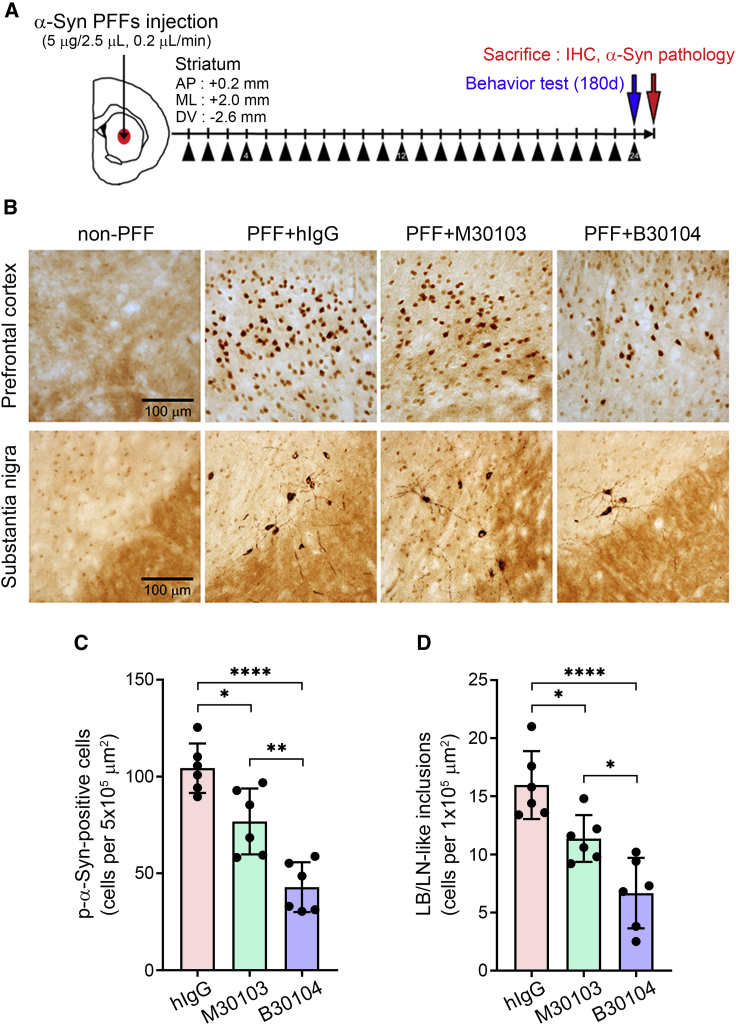
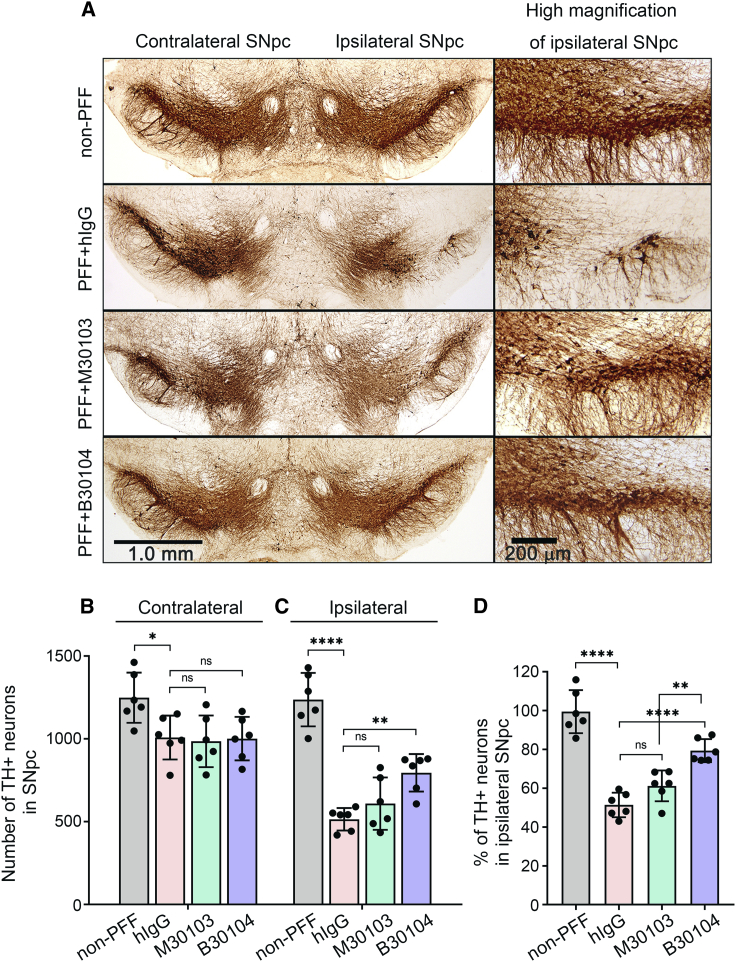
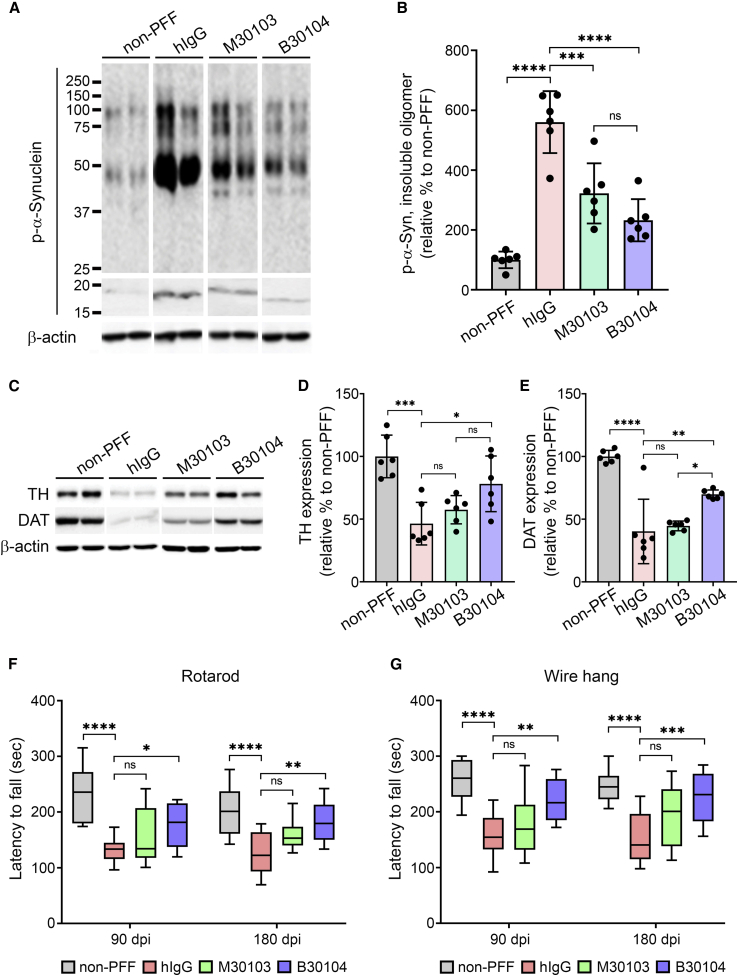
Effective delivery of therapeutics to the brain is challenging. Molecular shuttles use receptors expressed on brain endothelial cells to deliver therapeutics. Antibodies targeting transferrin receptor (TfR) have been widely developed as molecular shuttles. However, the TfR-based approach raises concerns about safety and developmental burden. Here, we report insulin-like growth factor 1 receptor (IGF1R) as an ideal target for the molecular shuttle. We also describe Grabody B, an antibody against IGF1R, as a molecular shuttle. Grabody B has broad cross-species reactivity and does not interfere with IGF1R-mediated signaling. We demonstrate that administration of Grabody B-fused anti-alpha-synuclein (α-Syn) antibody induces better improvement in neuropathology and behavior in a Parkinson's disease animal model than the therapeutic antibody alone due to its superior serum pharmacokinetics and enhanced brain exposure. The results indicate that IGF1R is an ideal shuttle target and Grabody B is a safe and efficient molecular shuttle.
Keywords: Grabody B; Parkinson’s disease; alpha-synuclein; blood-brain barrier; brain endothelial cells; insulin-like growth factor 1 receptor; molecular shuttle; neurodegenerative diseases; pre-formed fibrils; transferrin receptor.
© 2022 The Author(s).
Conflict of interest statement
Authors affiliated with ABL Bio are employees with stakes in the company. The work by J.-W.S., A.S.H., and D.B.S., and the work by S.M. and J.L. described in the manuscript, was funded by ABL Bio.
Figures
References
-
- Boado R.J., Zhang Y., Zhang Y., Pardridge W.M. Humanization of anti-human insulin receptor antibody for drug targeting across the human blood–brain barrier. Biotechnol. Bioeng. 2007;96:381–391. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
