

Cutting Edge: Neutrophil Complement Receptor Signaling Is Required for BAFF-Dependent Humoral Responses in Mice
- PMID: 36454023
- PMCID: PMC9780177
- DOI: 10.4049/jimmunol.2200410
Cutting Edge: Neutrophil Complement Receptor Signaling Is Required for BAFF-Dependent Humoral Responses in Mice
Abstract
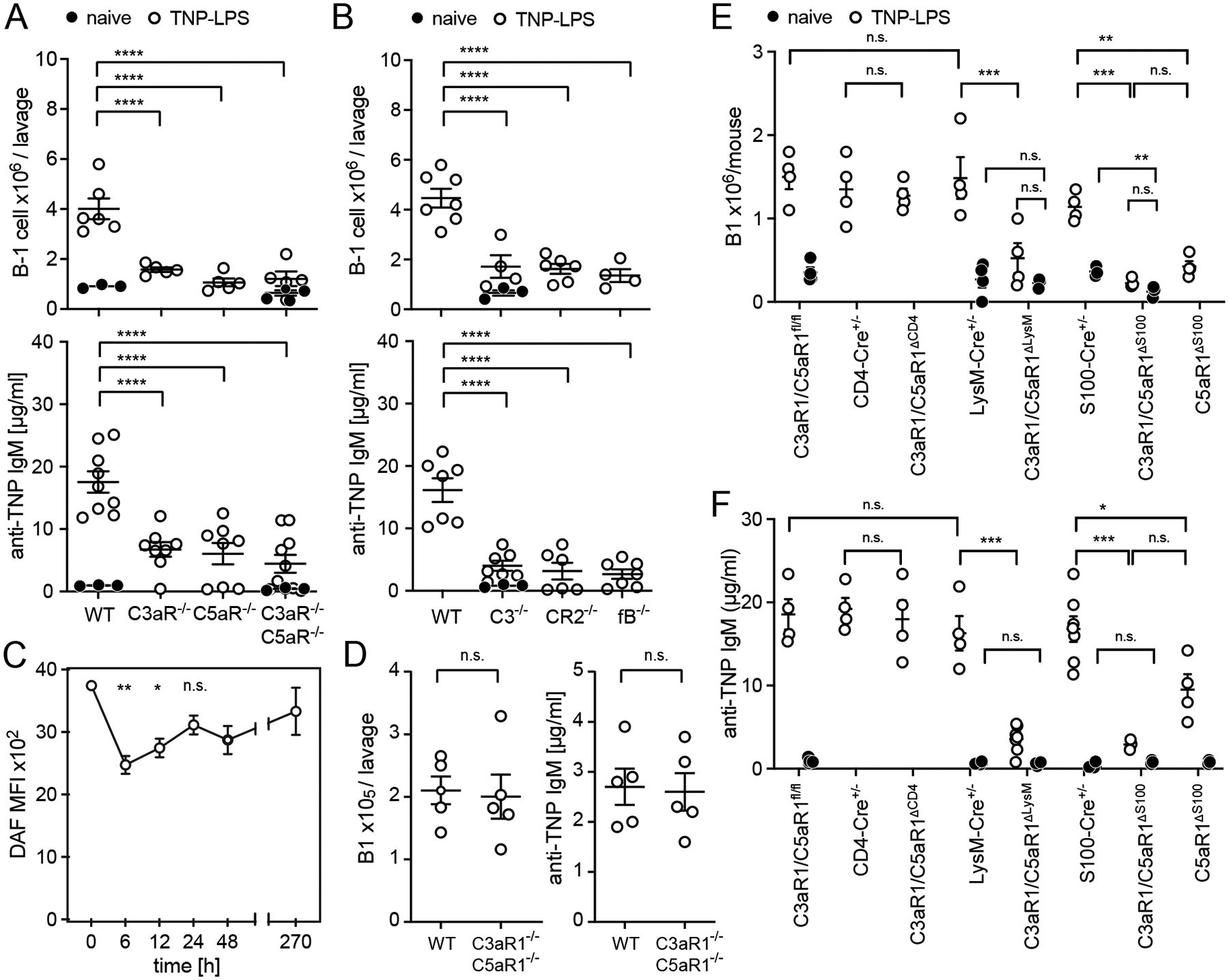
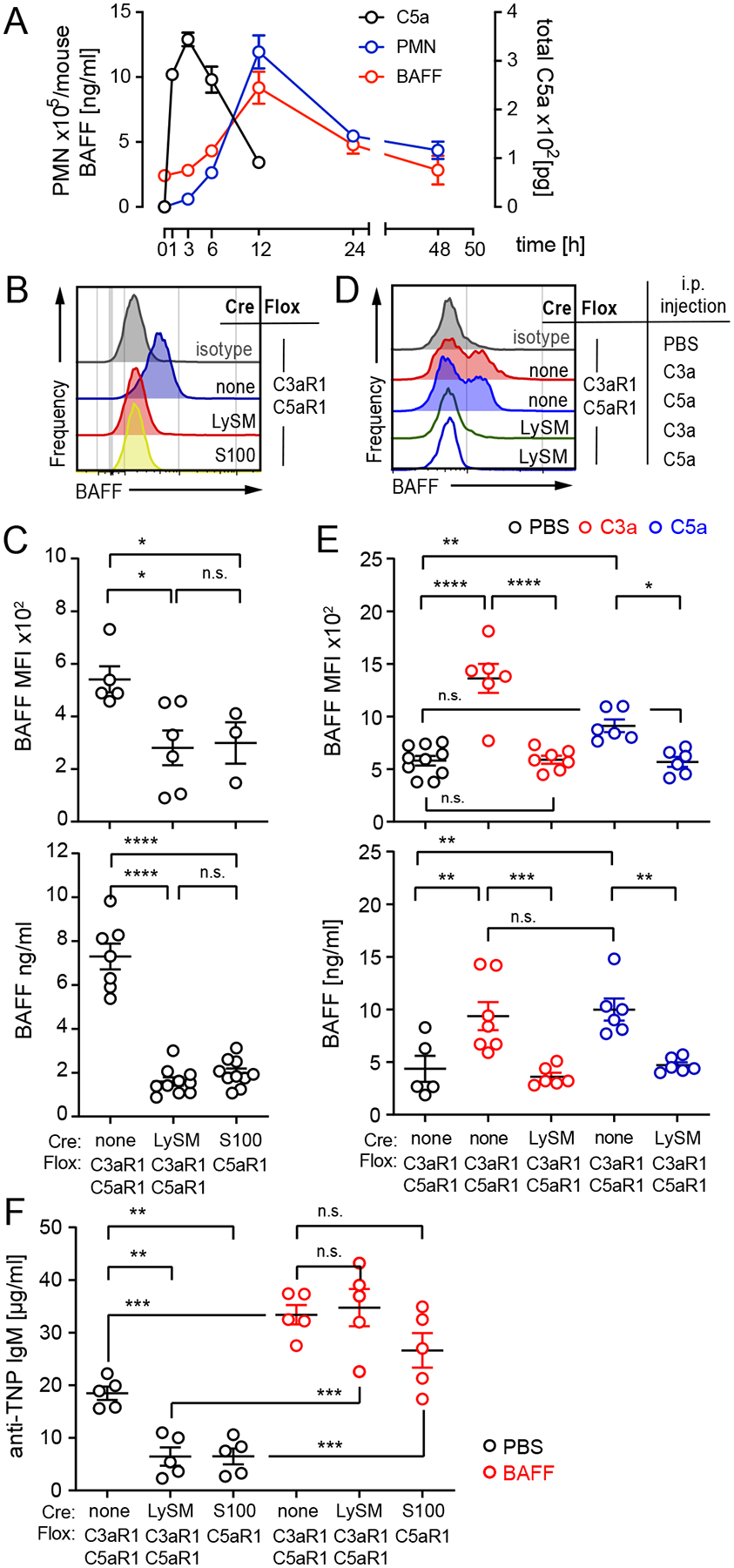
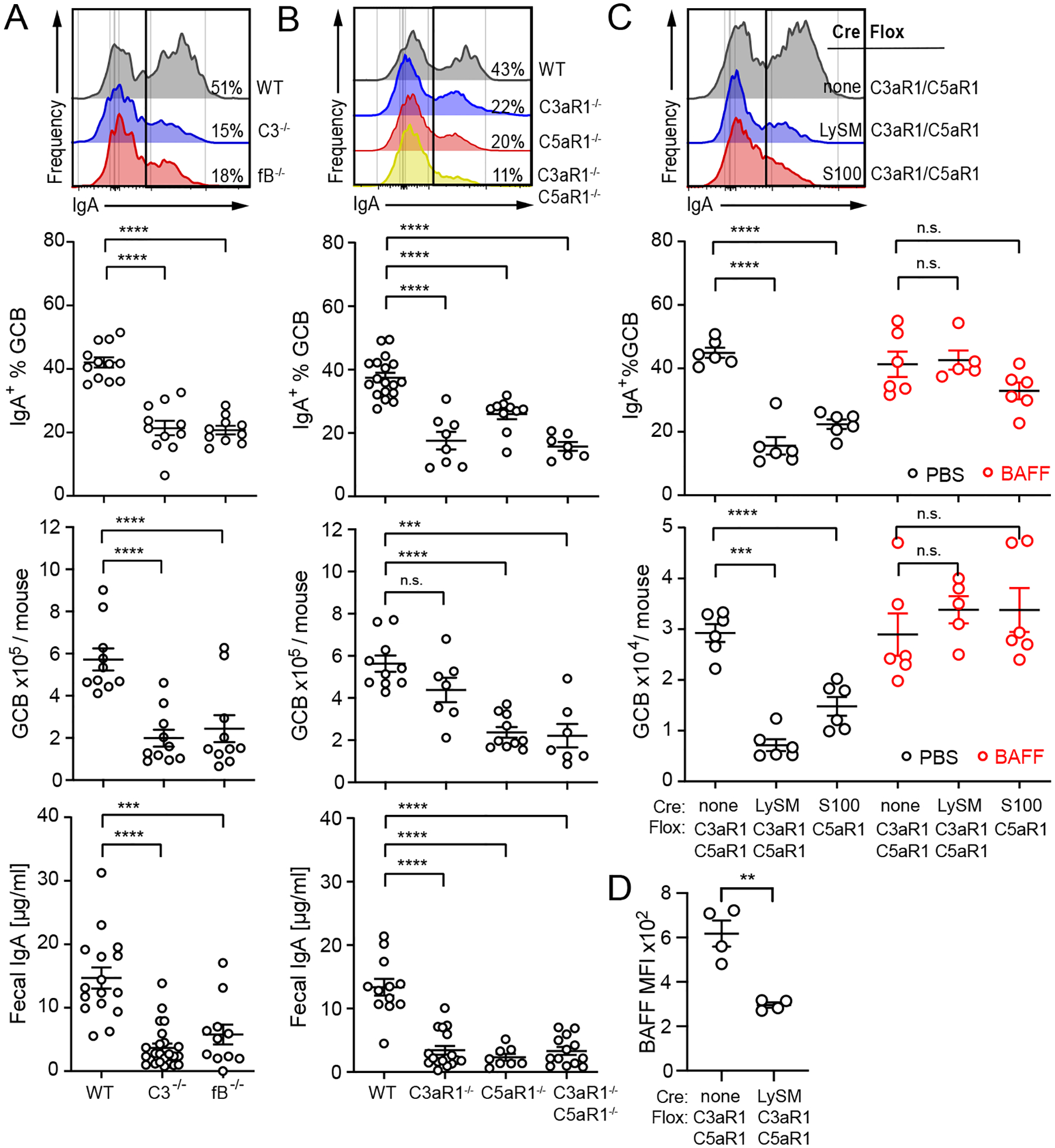
T cell-independent (TI) B cell responses to nonprotein Ags involve multiple cues from the innate immune system. Neutrophils express complement receptors and activated neutrophils can release BAFF, but mechanisms effectively linking neutrophil activation to TI B cell responses are incompletely understood. Using germline and conditional knockout mice, we found that TI humoral responses involve alternative pathway complement activation and neutrophil-expressed C3a and C5a receptors (C3aR1/C5aR1) that promote BAFF-dependent B1 cell expansion and TI Ab production. Conditional absence of C3aR1/C5aR1 on neutrophils lowered serum BAFF levels, led to fewer Peyer's patch germinal center B cells, reduced germinal center B cells IgA class-switching, and lowered fecal IgA levels. Together, the results indicate that sequential activation of complement on neutrophils crucially supports humoral TI and mucosal IgA responses through upregulating neutrophil production of BAFF.
Copyright © 2022 by The American Association of Immunologists, Inc.
Figures
Similar articles
-
Follicular B2 Cell Activation and Class Switch Recombination Depend on Autocrine C3ar1/C5ar1 Signaling in B2 Cells.J Immunol. 2019 Jul 15;203(2):379-388. doi: 10.4049/jimmunol.1900276. Epub 2019 Jun 19. J Immunol. 2019. PMID: 31217324 Free PMC article.
-
Neutrophils contribute to excess serum BAFF levels and promote CD4+ T cell and B cell responses in lupus-prone mice.PLoS One. 2014 Jul 10;9(7):e102284. doi: 10.1371/journal.pone.0102284. eCollection 2014. PLoS One. 2014. PMID: 25010693 Free PMC article.
-
Complement receptor C3aR1 controls neutrophil mobilization following spinal cord injury through physiological antagonism of CXCR2.JCI Insight. 2019 May 2;4(9):e98254. doi: 10.1172/jci.insight.98254. eCollection 2019 May 2. JCI Insight. 2019. PMID: 31045582 Free PMC article.
-
Re-thinking the functions of IgA(+) plasma cells.Gut Microbes. 2014;5(5):652-62. doi: 10.4161/19490976.2014.969977. Gut Microbes. 2014. PMID: 25483334 Free PMC article. Review.
-
Mucosal immunoregulation: environmental lipopolysaccharide and GALT T lymphocytes regulate the IgA response.Microbiol Immunol. 1984;28(3):261-80. doi: 10.1111/j.1348-0421.1984.tb00679.x. Microbiol Immunol. 1984. PMID: 6234450 Review.
Cited by
-
Translating B cell immunology to the treatment of antibody-mediated allograft rejection.Nat Rev Nephrol. 2024 Apr;20(4):218-232. doi: 10.1038/s41581-023-00791-0. Epub 2024 Jan 2. Nat Rev Nephrol. 2024. PMID: 38168662 Review.
-
Current understanding of the molecular mechanisms of circulating permeability factor in focal segmental glomerulosclerosis.Front Immunol. 2023 Sep 19;14:1247606. doi: 10.3389/fimmu.2023.1247606. eCollection 2023. Front Immunol. 2023. PMID: 37795085 Free PMC article. Review.
-
How Neutrophils Shape the Immune Response: Reassessing Their Multifaceted Role in Health and Disease.Int J Mol Sci. 2023 Dec 18;24(24):17583. doi: 10.3390/ijms242417583. Int J Mol Sci. 2023. PMID: 38139412 Free PMC article. Review.
References
-
- Ahearn JM, Fischer MB, Croix D, Goerg S, Ma M, Xia J, Zhou X, Howard RG, Rothstein TL, and Carroll MC. 1996. Disruption of the Cr2 locus results in a reduction in B-1a cells and in an impaired B cell response to T-dependent antigen. Immunity 4: 251–262. - PubMed
-
- Dempsey PW, Allison ME, Akkaraju S, Goodnow CC, and Fearon DT. 1996. C3d of complement as a molecular adjuvant: bridging innate and acquired immunity. Science 271: 348–350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
