

AKT-driven epithelial-mesenchymal transition is affected by copper bioavailability in HER2 negative breast cancer cells via a LOXL2-independent mechanism
- PMID: 36454513
- PMCID: PMC9947069
- DOI: 10.1007/s13402-022-00738-w
AKT-driven epithelial-mesenchymal transition is affected by copper bioavailability in HER2 negative breast cancer cells via a LOXL2-independent mechanism
Abstract
Background: The main mechanism underlying cancer dissemination is the epithelial to mesenchymal transition (EMT). This process is orchestrated by cytokines like TGFβ, involving "non-canonical" AKT- or STAT3-driven pathways. Recently, the alteration of copper homeostasis seems involved in the onset and progression of cancer.
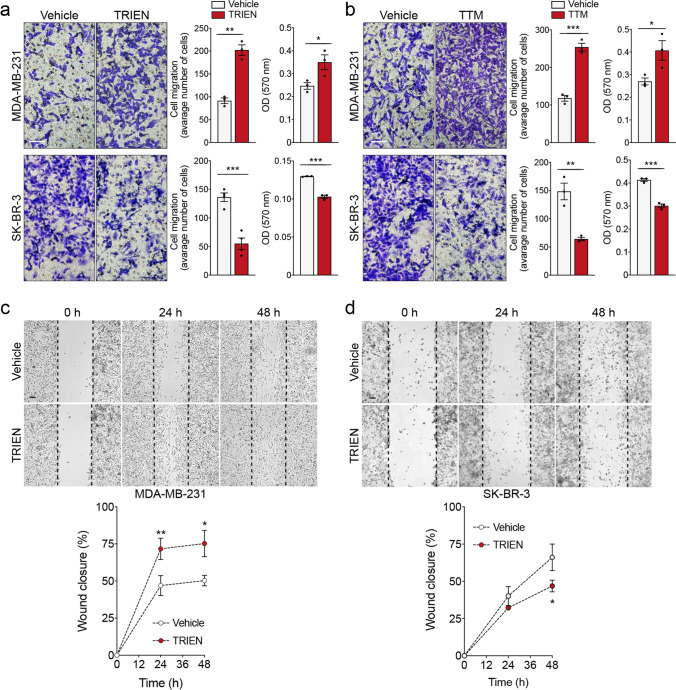
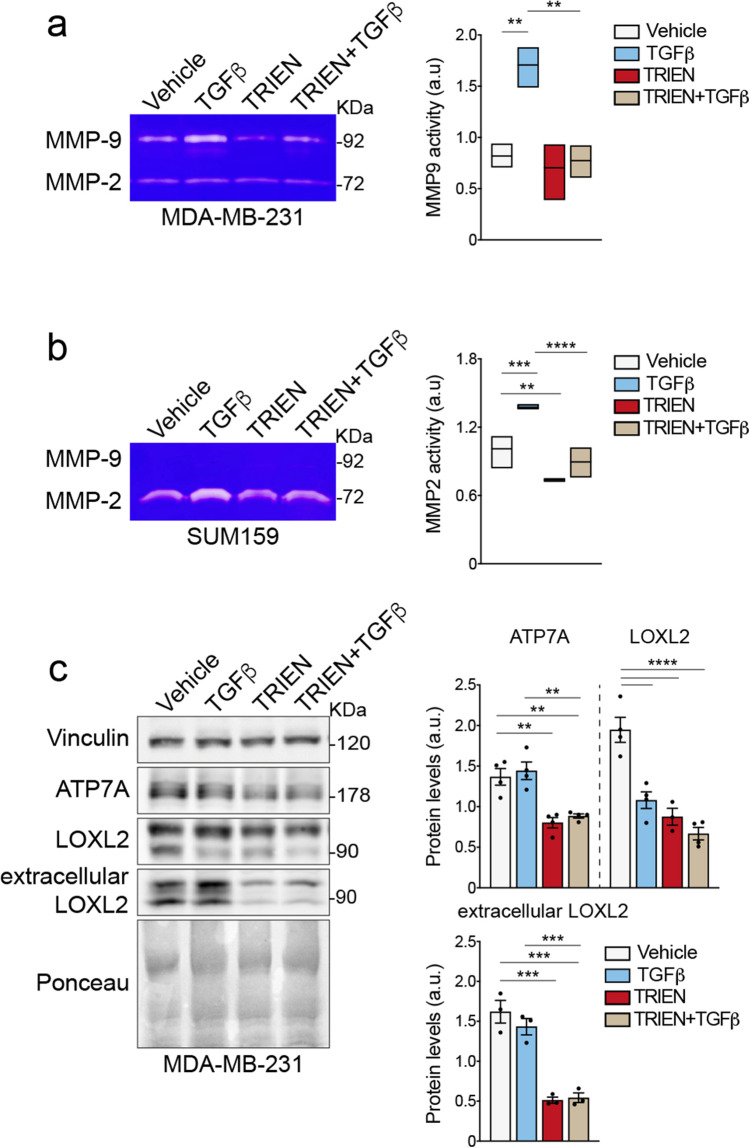
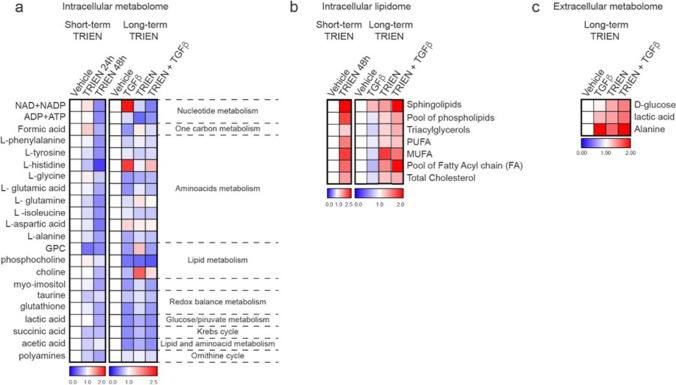
Methods: We expose different breast cancer cell lines, including two triple negative (TNBC) ones, an HER2 enriched and one cell line representative of the Luminal A molecular subtype, to short- or long-term copper-chelation by triethylenetetramine (TRIEN). We analyse changes in the expression of EMT markers (E-cadherin, fibronectin, vimentin and αSMA), in the levels and activity of extracellular matrix components (LOXL2, fibronectin and MMP2/9) and of copper homeostasis markers by Western blot analyses, immunofluorescence, enzyme activity assays and RT-qPCR. Boyden Chamber and wound healing assays revealed the impact of copper chelation on cell migration. Additionally, we explored whether perturbation of copper homeostasis affects EMT prompted by TGFβ. Metabolomic and lipidomic analyses were applied to search the effects of copper chelation on the metabolism of breast cancer cells. Finally, bioinformatics analysis of data on breast cancer patients obtained from different databases was employed to correlate changes in kinases and copper markers with patients' survival.
Results: Remarkably, only HER2 negative breast cancer cells differently responded to short- or long-term exposure to TRIEN, initially becoming more aggressive but, upon prolonged exposure, retrieving epithelial features, reducing their invasiveness. This phenomenon may be related to the different impact of the short and prolonged activation of the AKT kinase and to the repression of STAT3 signalling. Bioinformatics analyses confirmed the positive correlation of breast cancer patients' survival with AKT activation and up-regulation of CCS. Eventually, metabolomics studies demonstrate a prevalence of glycolysis over mitochondrial energetic metabolism and of lipidome changes in TNBC cells upon TRIEN treatment.
Conclusions: We provide evidence of a pivotal role of copper in AKT-driven EMT activation, acting independently of HER2 in TNBC cells and via a profound change in their metabolism. Our results support the use of copper-chelators as an adjuvant therapeutic strategy for TNBC.
Keywords: AKT; Breast cancer; Copper; Epithelial to mesenchymal transition; HER2; LOXL2; TGFβ; TRIEN.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
SHP2 promotes the epithelial-mesenchymal transition in triple negative breast cancer cells by regulating β-catenin.J Cancer Res Clin Oncol. 2025 Jan 29;151(2):55. doi: 10.1007/s00432-025-06097-x. J Cancer Res Clin Oncol. 2025. PMID: 39878804 Free PMC article.
-
Tumor-associated macrophages promote epithelial-mesenchymal transition and the cancer stem cell properties in triple-negative breast cancer through CCL2/AKT/β-catenin signaling.Cell Commun Signal. 2022 Jun 17;20(1):92. doi: 10.1186/s12964-022-00888-2. Cell Commun Signal. 2022. PMID: 35715860 Free PMC article.
-
Elevated TGF-β1 and -β2 expression accelerates the epithelial to mesenchymal transition in triple-negative breast cancer cells.Cytokine. 2015 Sep;75(1):151-8. doi: 10.1016/j.cyto.2015.05.020. Epub 2015 Jun 15. Cytokine. 2015. PMID: 26088755
-
Triple-Negative Breast Cancer Progression and Drug Resistance in the Context of Epithelial-Mesenchymal Transition.Cancers (Basel). 2025 Jan 12;17(2):228. doi: 10.3390/cancers17020228. Cancers (Basel). 2025. PMID: 39858010 Free PMC article. Review.
-
TGFβ-induced metabolic reprogramming during epithelial-to-mesenchymal transition in cancer.Cell Mol Life Sci. 2020 Jun;77(11):2103-2123. doi: 10.1007/s00018-019-03398-6. Epub 2019 Dec 10. Cell Mol Life Sci. 2020. PMID: 31822964 Free PMC article. Review.
Cited by
-
Copper's new role in cancer: how cuproptosis-related genes could revolutionize glioma treatment.BMC Cancer. 2025 May 12;25(1):859. doi: 10.1186/s12885-025-14151-7. BMC Cancer. 2025. PMID: 40355831 Free PMC article.
-
Exploring lipidomic profiles and their correlation with hormone receptor and HER2 status in breast cancer.Oncol Lett. 2024 Oct 29;29(1):34. doi: 10.3892/ol.2024.14781. eCollection 2025 Jan. Oncol Lett. 2024. PMID: 39512509 Free PMC article.
-
BET inhibitors (BETi) influence oxidative phosphorylation metabolism by affecting mitochondrial dynamics leading to alterations in apoptotic pathways in triple-negative breast cancer (TNBC) cells.Cell Prolif. 2024 Dec;57(12):e13730. doi: 10.1111/cpr.13730. Epub 2024 Sep 2. Cell Prolif. 2024. PMID: 39223828 Free PMC article.
-
Roles of Copper Transport Systems Members in Breast Cancer.Cancer Med. 2024 Dec;13(24):e70498. doi: 10.1002/cam4.70498. Cancer Med. 2024. PMID: 39676279 Free PMC article. Review.
-
Characterization of tumor prognosis and sensitive chemotherapy drugs based on cuproptosis-related gene signature in ovarian cancer.BMC Womens Health. 2025 Jan 24;25(1):37. doi: 10.1186/s12905-024-03519-9. BMC Womens Health. 2025. PMID: 39849417 Free PMC article.
References
-
- A. De Luca, A. Barile, M. Arciello, L. Rossi, Copper Homeostasis as Target of Both Consolidated and Innovative Strategies of Anti-Tumor Therapy. J. Trace Elem. Med. Biol. 55 (2019). 10.1016/j.jtemb.2019.06.008 - PubMed
-
- Deshmukh AP, Vasaikar SV, Tomczak K, Tripathi S, Den Hollander P, Arslan E, Chakraborty P, Soundararajan R, Jolly MK, Rai K, et al. Identification of EMT Signaling Cross-Talk and Gene Regulatory Networks by Single-Cell RNA Sequencing. Proc. Natl. Acad. Sci. U. S. A. 2021;118:2102050118. doi: 10.1073/pnas.2102050118. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
