

Dysregulated long non-coding RNA in Sjögren's disease impacts both interferon and adaptive immune responses
- PMID: 36456101
- PMCID: PMC9717416
- DOI: 10.1136/rmdopen-2022-002672
Dysregulated long non-coding RNA in Sjögren's disease impacts both interferon and adaptive immune responses
Abstract
Objective: Sjögren's disease (SjD) is an autoimmune disease characterised by inflammatory destruction of exocrine glands. Patients with autoantibodies to Ro/SSA (SjDRo+) exhibit more severe disease. Long non-coding RNAs (lncRNAs) are a functionally diverse class of non-protein-coding RNAs whose role in autoimmune disease pathology has not been well characterised.
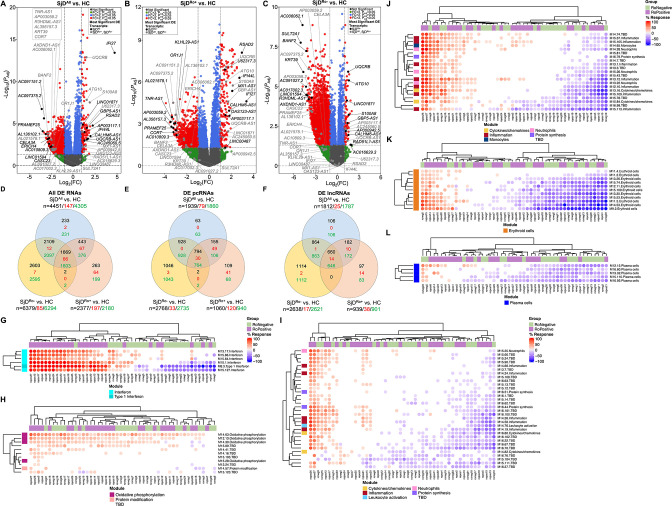
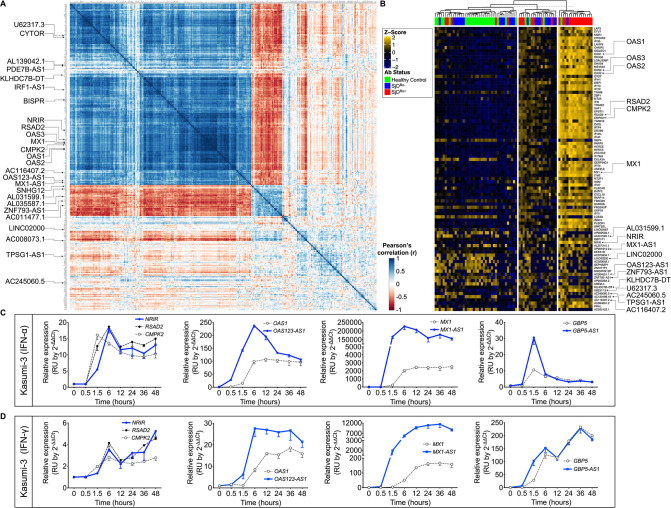
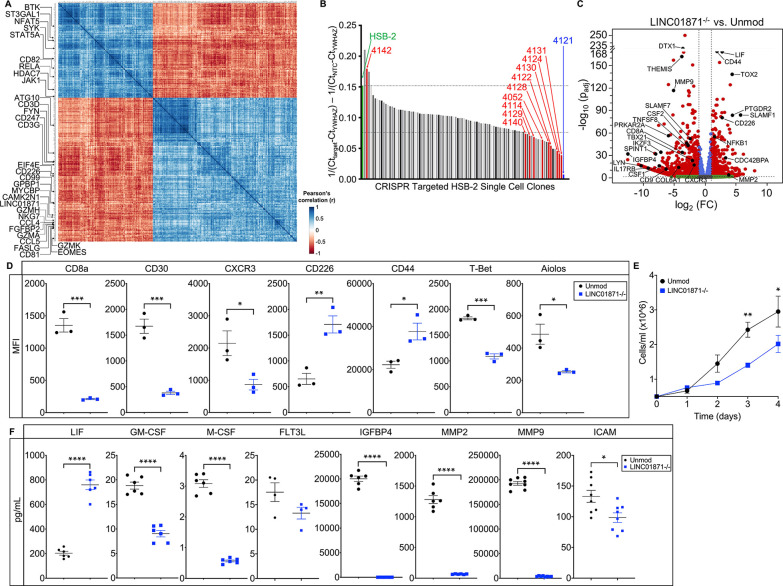
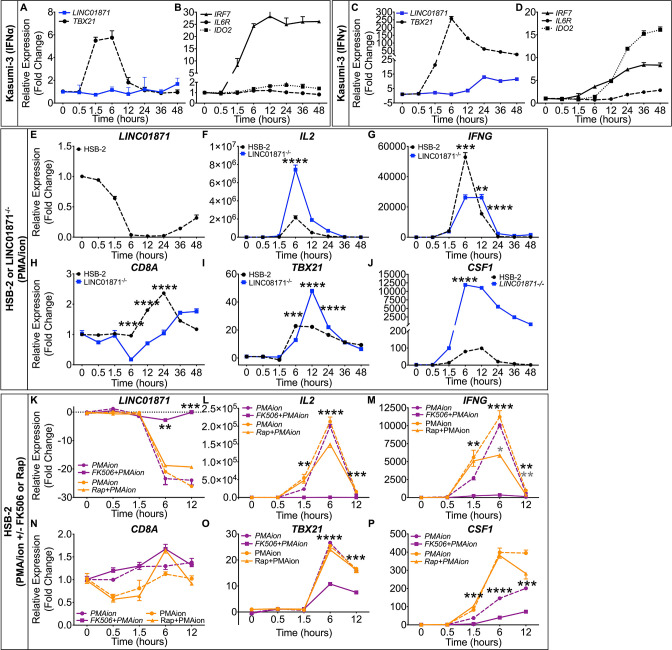
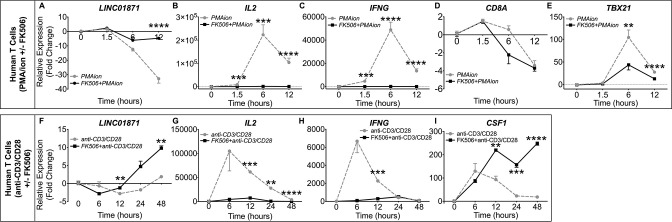
Methods: Whole blood RNA-sequencing (RNA-seq) was performed on SjD cases (n=23 Ro/SSA negative (SjDRo-); n=27 Ro/SSA positive (SjDRo+) and healthy controls (HCs; n=27). Bioinformatics and pathway analyses of differentially expressed (DE) transcripts (log2 fold change ≥2 or ≤0.5; padj<0.05) were used to predict lncRNA function. LINC01871 was characterised by RNA-seq analyses of HSB-2 cells with CRISPR-targeted LINC01871 deletion (LINC01871-/ -) and in vitro stimulation assays.
Results: Whole blood RNA-seq revealed autoantibody-specific transcription profiles and disproportionate downregulation of DE transcripts in SjD cases relative to HCs. Sixteen DE lncRNAs exhibited correlated expression with the interferon (IFN)-regulated gene, RSAD2, in SjDRo+ (r≥0.65 or ≤-0.6); four antisense lncRNAs exhibited IFN-regulated expression in immune cell lines. LINC01871 was upregulated in all SjD cases. RNA-seq and pathway analyses of LINC01871-/ - cells implicated roles in cytotoxic function, differentiation and IFNγ induction. LINC01871 was induced by IFNγ in a myeloid cell line and regulated by calcineurin/NFAT pathway and T cell receptor (TCR) signalling in primary human T cells.
Conclusion: LINC01871 influences expression of many immune cell genes and growth factors, is IFNγ inducible, and regulated by calcineurin signalling and TCR ligand engagement. Altered LINC01871 expression may influence the dysregulated T cell inflammatory pathways implicated in SjD.
Trial registration: ClinicalTrials.gov NCT02327884.
Keywords: Autoantibodies; Autoimmune Diseases; Autoimmunity; Polymorphism, Genetic; Sjogren's Syndrome.
© Author(s) (or their employer(s)) 2022. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: KS is a current employee of Janssen. ADF and CJL have an active collaborative research agreement with Janssen. All other authors have reported that they have no competing interests to report.
Figures
Comment in
-
Interferon signature in systemic autoimmune diseases: what does it mean?RMD Open. 2022 Dec;8(2):e002687. doi: 10.1136/rmdopen-2022-002687. RMD Open. 2022. PMID: 36597991 Free PMC article. No abstract available.
Similar articles
-
Identification of novel autoantibodies in Sjögren's disease.Front Immunol. 2025 Feb 3;16:1524940. doi: 10.3389/fimmu.2025.1524940. eCollection 2025. Front Immunol. 2025. PMID: 39963128 Free PMC article.
-
Transcriptomic Profiling of Peripheral B Cells in Antibody Positive Sjogren's Patients Reveals Interferon Signature.Genes (Basel). 2024 May 15;15(5):628. doi: 10.3390/genes15050628. Genes (Basel). 2024. PMID: 38790257 Free PMC article.
-
Long Non-Coding RNAs in Sjögren's Disease.Int J Mol Sci. 2024 May 9;25(10):5162. doi: 10.3390/ijms25105162. Int J Mol Sci. 2024. PMID: 38791207 Free PMC article. Review.
-
Inhibition of JAK-STAT pathway corrects salivary gland inflammation and interferon driven immune activation in Sjögren's disease.Ann Rheum Dis. 2024 Jul 15;83(8):1034-1047. doi: 10.1136/ard-2023-224842. Ann Rheum Dis. 2024. PMID: 38527764 Free PMC article.
-
Is minor salivary gland biopsy still mandatory in Sjogren's syndrome? Does seronegative Sjogren's syndrome exist?Autoimmun Rev. 2024 Jan;23(1):103425. doi: 10.1016/j.autrev.2023.103425. Epub 2023 Aug 25. Autoimmun Rev. 2024. PMID: 37634677 Review.
Cited by
-
Multiomic single-cell sequencing defines tissue-specific responses in Stevens-Johnson Syndrome and Toxic epidermal necrolysis.bioRxiv [Preprint]. 2024 Feb 18:2023.11.26.568771. doi: 10.1101/2023.11.26.568771. bioRxiv. 2024. Update in: Nat Commun. 2024 Oct 8;15(1):8722. doi: 10.1038/s41467-024-52990-3. PMID: 38405793 Free PMC article. Updated. Preprint.
-
Variants in the DDX6-CXCR5 autoimmune disease risk locus influence the regulatory network in immune cells and salivary gland.bioRxiv [Preprint]. 2023 Oct 6:2023.10.05.561076. doi: 10.1101/2023.10.05.561076. bioRxiv. 2023. Update in: Ann Rheum Dis. 2025 May 29:S0003-4967(25)00949-5. doi: 10.1016/j.ard.2025.04.023. PMID: 39071447 Free PMC article. Updated. Preprint.
-
Role of ncRNAs in the Pathogenesis of Sjögren's Syndrome.Biomedicines. 2024 Jul 11;12(7):1540. doi: 10.3390/biomedicines12071540. Biomedicines. 2024. PMID: 39062113 Free PMC article. Review.
-
[Sex-specific aspects in rheumatology].Z Rheumatol. 2024 Nov;83(9):749-759. doi: 10.1007/s00393-024-01527-6. Epub 2024 Jun 13. Z Rheumatol. 2024. PMID: 38869669 German.
-
Multiomic single-cell sequencing defines tissue-specific responses in Stevens-Johnson syndrome and toxic epidermal necrolysis.Nat Commun. 2024 Oct 8;15(1):8722. doi: 10.1038/s41467-024-52990-3. Nat Commun. 2024. PMID: 39379371 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- U54 GM104938/GM/NIGMS NIH HHS/United States
- R21 AR079089/AR/NIAMS NIH HHS/United States
- R01 AR065953/AR/NIAMS NIH HHS/United States
- R01 AR073855/AR/NIAMS NIH HHS/United States
- R33 AR076803/AR/NIAMS NIH HHS/United States
- R01 AR074310/AR/NIAMS NIH HHS/United States
- Z01 DE000704/ImNIH/Intramural NIH HHS/United States
- P20 GM103456/GM/NIGMS NIH HHS/United States
- P50 AR060804/AR/NIAMS NIH HHS/United States
- UM1 AI144292/AI/NIAID NIH HHS/United States
- R01 HL113326/HL/NHLBI NIH HHS/United States
- P30 AR073750/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials