

Yeast-based evolutionary modeling of androgen receptor mutations and natural selection
- PMID: 36459502
- PMCID: PMC9718406
- DOI: 10.1371/journal.pgen.1010518
Yeast-based evolutionary modeling of androgen receptor mutations and natural selection
Abstract
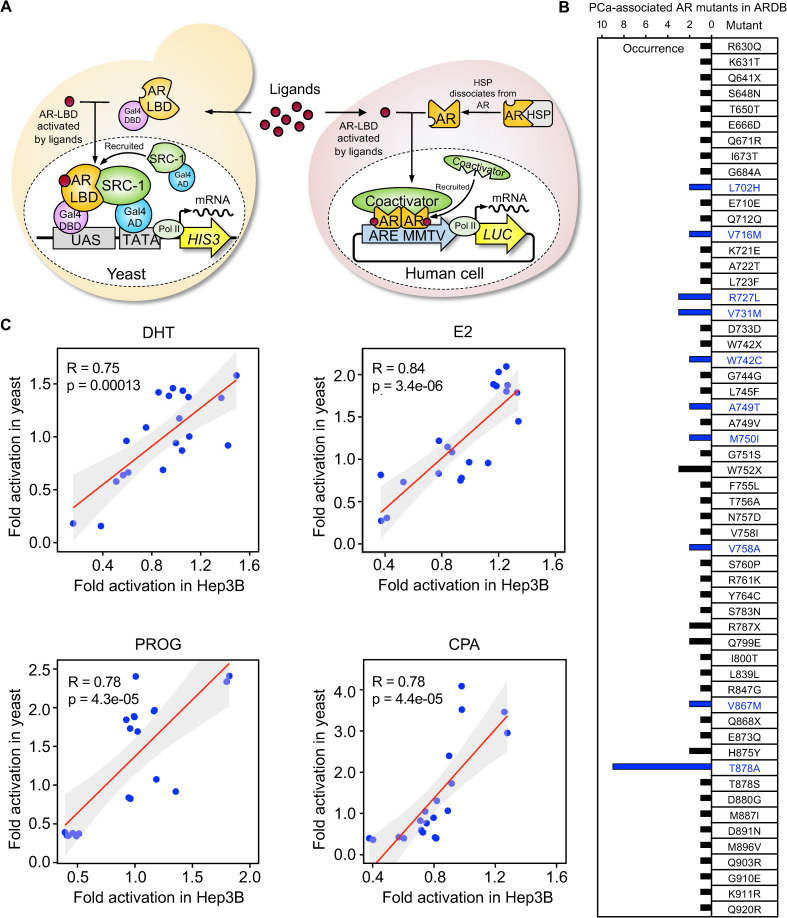
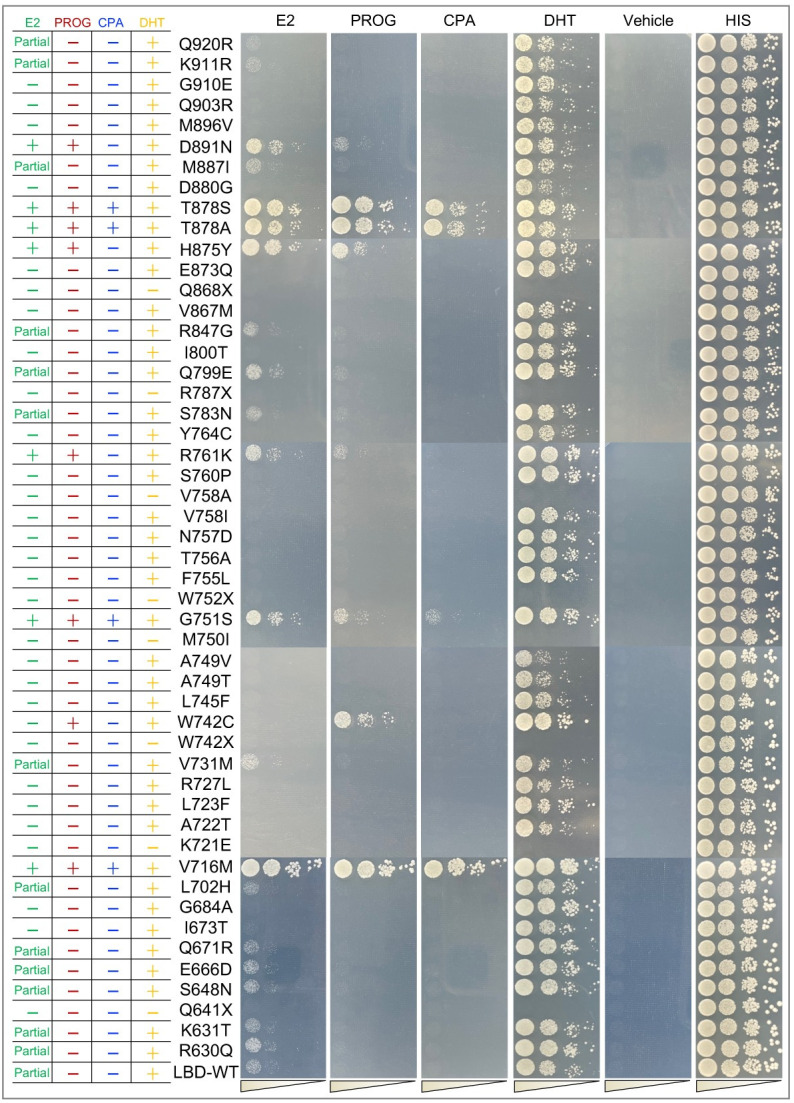
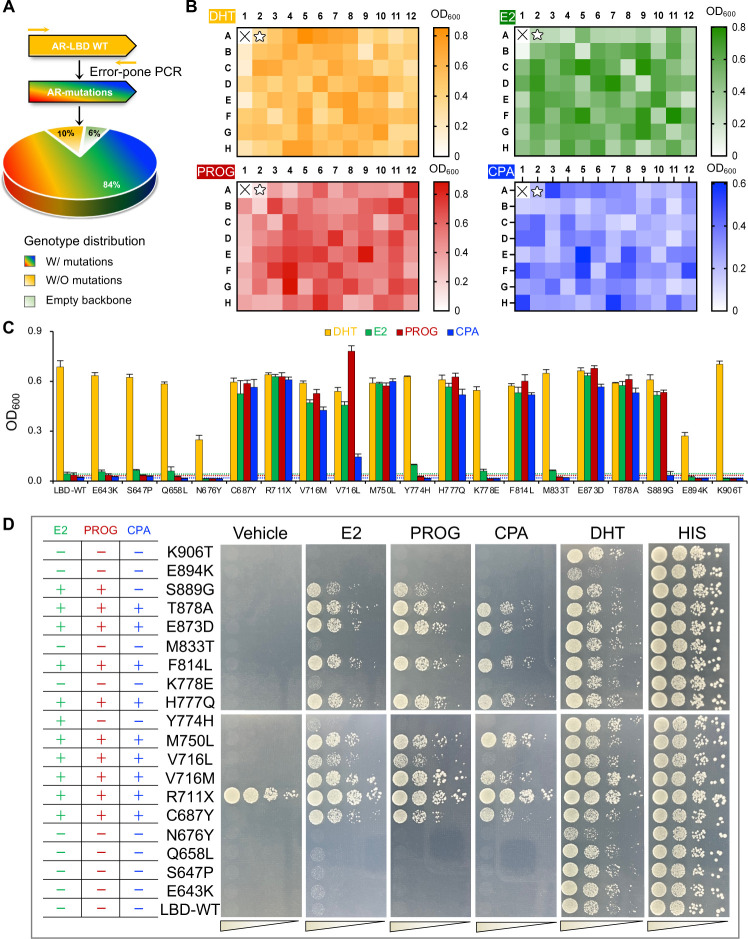
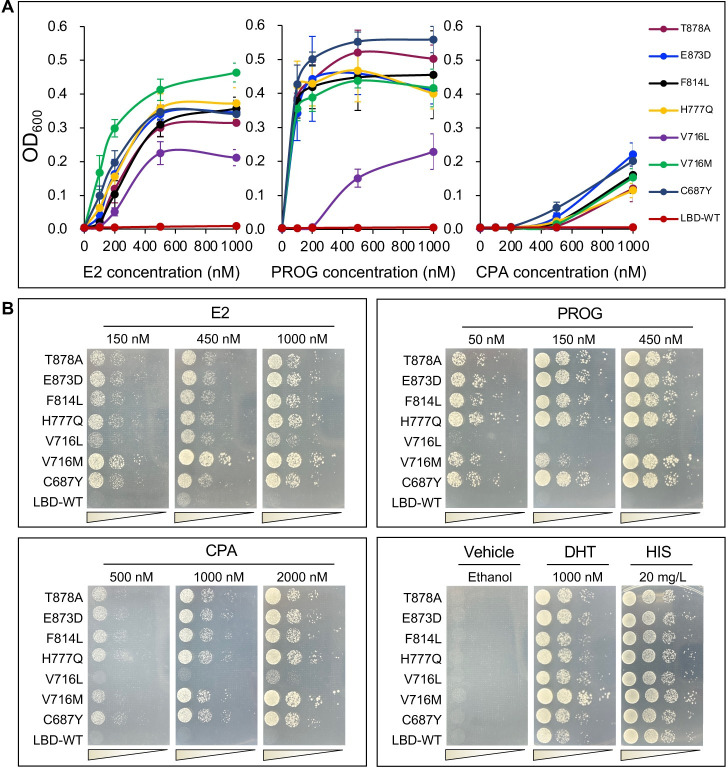
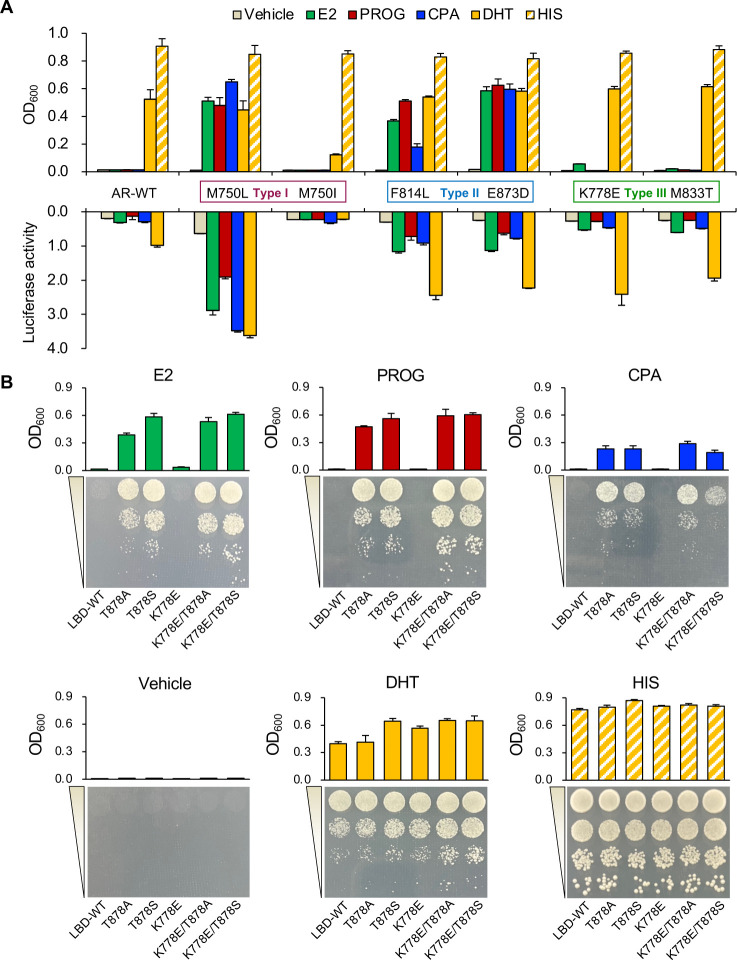
Cancer progression is associated with the evolutionary accumulation of genetic mutations that are biologically significant. Mutations of the androgen receptor (AR) are associated with the development of prostate cancer (PCa) by responding to non-androgenic hormones, and the lack of annotations in their responsiveness to hormone ligands remains a daunting challenge. Here, we have used a yeast reporter system to quickly evaluate the responsiveness of all fifty clinical AR mutations to a variety of steroidal ligands including dihydrotestosterone (DHT), 17β-estradiol (E2), progesterone (PROG), and cyproterone acetate (CPA). Based on an AR-driven reporter that synthesizes histidine, a basic amino acid required for yeast survival and propagation, the yeast reporter system enabling clonal selection was further empowered by combining with a random DNA mutagenesis library to simulate the natural evolution of AR gene under the selective pressures of steroidal ligands. In a time-frame of 1-2 weeks, 19 AR mutants were identified, in which 11 AR mutants were validated for activation by tested steroidal compounds. The high efficiency of our artificial evolution strategy was further evidenced by a sequential selection that enabled the discovery of multipoint AR mutations and evolution directions under the pressure of steroidal ligands. In summary, our designer yeast is a portable reporter module that can be readily adapted to streamline high-throughput AR-compound screening, used as a PCa clinical reference, and combined with additional bioassay systems to further extend its potential.
Copyright: © 2022 Zhang et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
There is potential Competing Interest. A patent has been filed for the screening method presented in this study.
Figures
Comment in
-
Modelling AR mutations.Nat Rev Urol. 2023 Feb;20(2):64. doi: 10.1038/s41585-022-00718-7. Nat Rev Urol. 2023. PMID: 36604519 No abstract available.
Similar articles
-
Simulating androgen receptor selection in designer yeast.Synth Syst Biotechnol. 2022 Aug 3;7(4):1108-1116. doi: 10.1016/j.synbio.2022.07.005. eCollection 2022 Dec. Synth Syst Biotechnol. 2022. PMID: 36017332 Free PMC article.
-
Isolation and characterization of the androgen receptor mutants with divergent transcriptional activity in response to hydroxyflutamide.Endocrine. 2000 Feb;12(1):69-76. doi: 10.1385/ENDO:12:1:69. Endocrine. 2000. PMID: 10855693
-
Activation of rat androgen receptor by androgenic ligands is unaffected by antiandrogens in Saccharomyces cerevisiae.Gene. 1998 Mar 16;209(1-2):247-54. doi: 10.1016/s0378-1119(98)00054-7. Gene. 1998. PMID: 9524277
-
Androgen-receptor gene structure and function in prostate cancer.World J Urol. 1996;14(5):329-37. doi: 10.1007/BF00184606. World J Urol. 1996. PMID: 8912473 Review.
-
Androgen receptor mutations in carcinoma of the prostate: significance for endocrine therapy.Am J Pharmacogenomics. 2001;1(4):241-9. doi: 10.2165/00129785-200101040-00001. Am J Pharmacogenomics. 2001. PMID: 12083956 Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
