

Decorating chromatin for enhanced genome editing using CRISPR-Cas9
- PMID: 36459645
- PMCID: PMC9894255
- DOI: 10.1073/pnas.2204259119
Decorating chromatin for enhanced genome editing using CRISPR-Cas9
Abstract
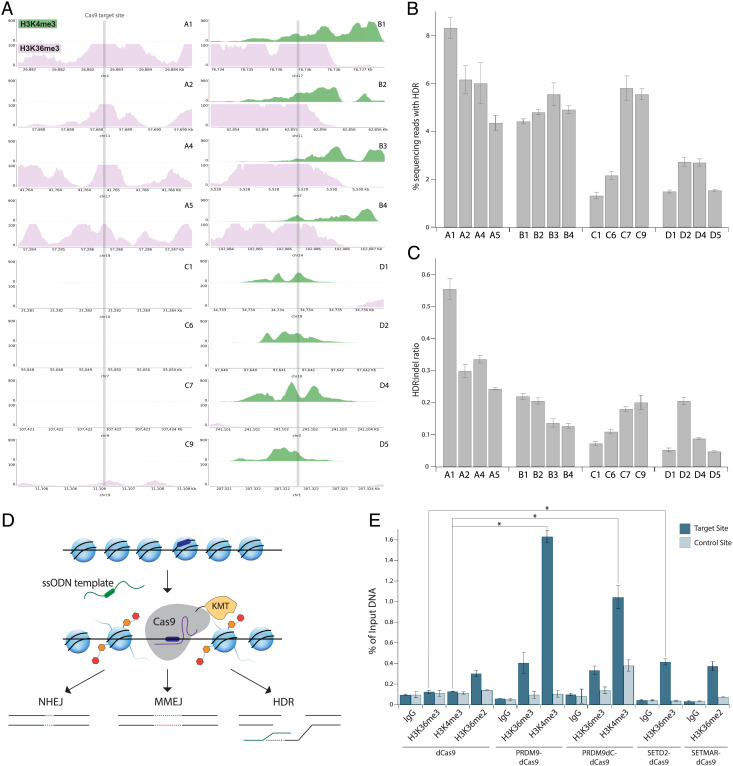
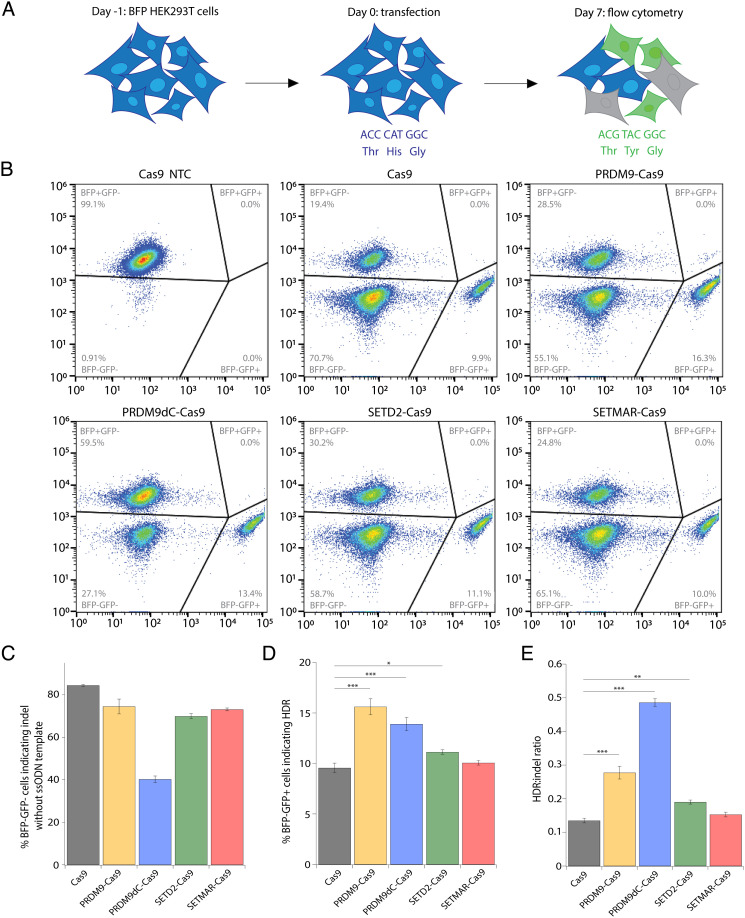
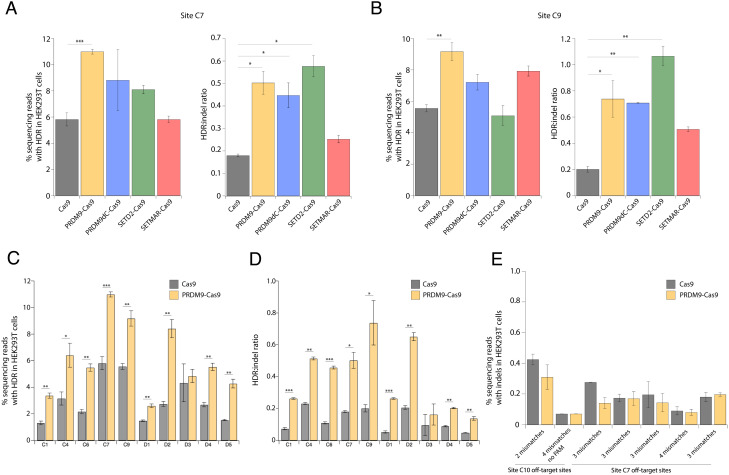
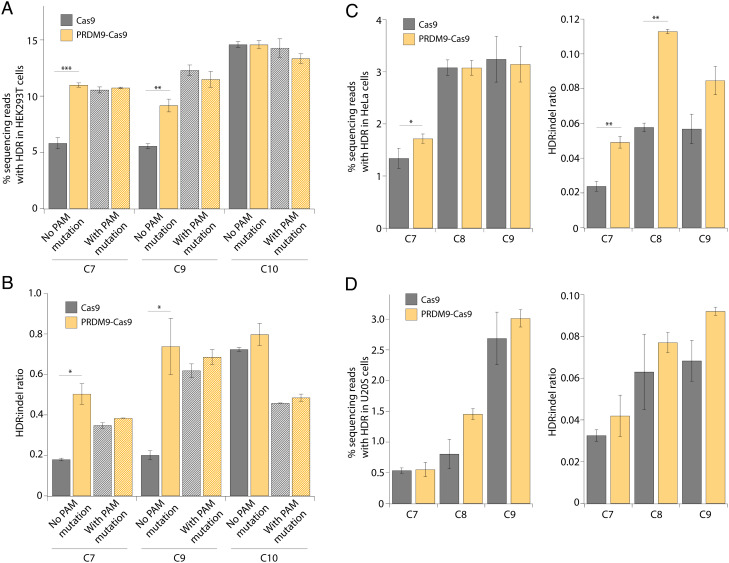
CRISPR-associated (Cas) enzymes have revolutionized biology by enabling RNA-guided genome editing. Homology-directed repair (HDR) in the presence of donor templates is currently the most versatile method to introduce precise edits following CRISPR-Cas-induced double-stranded DNA cuts, but HDR efficiency is generally low relative to end-joining pathways that lead to insertions and deletions (indels). We tested the hypothesis that HDR could be increased using a Cas9 construct fused to PRDM9, a chromatin remodeling factor that deposits histone methylations H3K36me3 and H3K4me3 to mediate homologous recombination in human cells. Our results show that the fusion protein contacts chromatin specifically at the Cas9 cut site in the genome to increase the observed HDR efficiency by threefold and HDR:indel ratio by fivefold compared with that induced by unmodified Cas9. HDR enhancement occurred in multiple cell lines with no increase in off-target genome editing. These findings underscore the importance of chromatin features for the balance between DNA repair mechanisms during CRISPR-Cas genome editing and provide a strategy to increase HDR efficiency.
Keywords: CRISPR; chromatin; epigenetics; genome editing.
Conflict of interest statement
The Regents of the University of California have patents issued and pending for CRISPR technologies on which J.A.D. is an inventor. J.A.D. is a cofounder of Caribou Biosciences, Editas Medicine, Scribe Therapeutics, Intellia Therapeutics, and Mammoth Biosciences. J.A.D. is a scientific advisory board member of Vertex, Caribou Biosciences, Intellia Therapeutics, Scribe Therapeutics, Mammoth Biosciences, Algen Biotechnologies, Felix Biosciences, The Column Group, and Inari. J.A.D. is Chief Science Advisor to Sixth Street, a Director at Johnson & Johnson, Altos and Tempus, and has research projects sponsored by Apple Tree Partners and Roche.
Figures
References
-
- Chapman J. R., Taylor M. R. G., Boulton S. J., Playing the end game: DNA double-strand break repair pathway choice. Mol. Cell 47, 497–510 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
