

Lactate-dependent chaperone-mediated autophagy induces oscillatory HIF-1α activity promoting proliferation of hypoxic cells
- PMID: 36462504
- PMCID: PMC10012408
- DOI: 10.1016/j.cels.2022.11.003
Lactate-dependent chaperone-mediated autophagy induces oscillatory HIF-1α activity promoting proliferation of hypoxic cells
Abstract
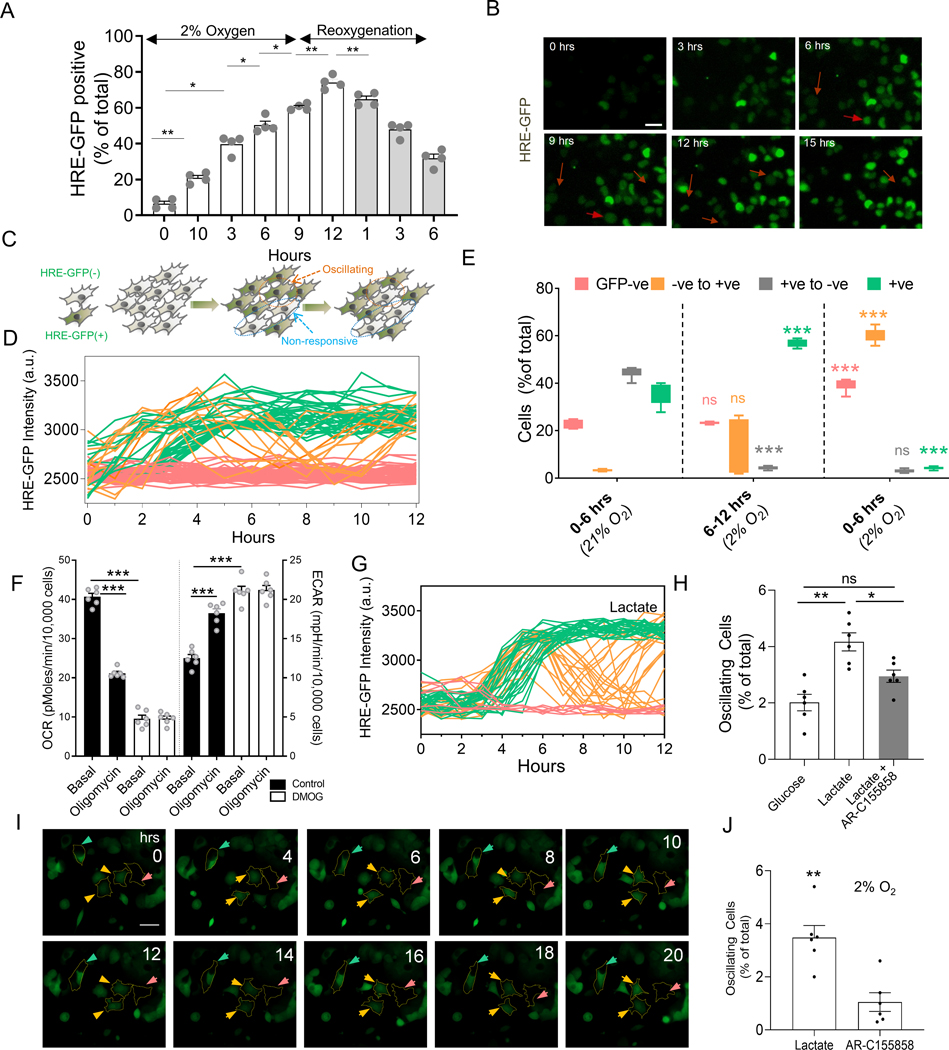
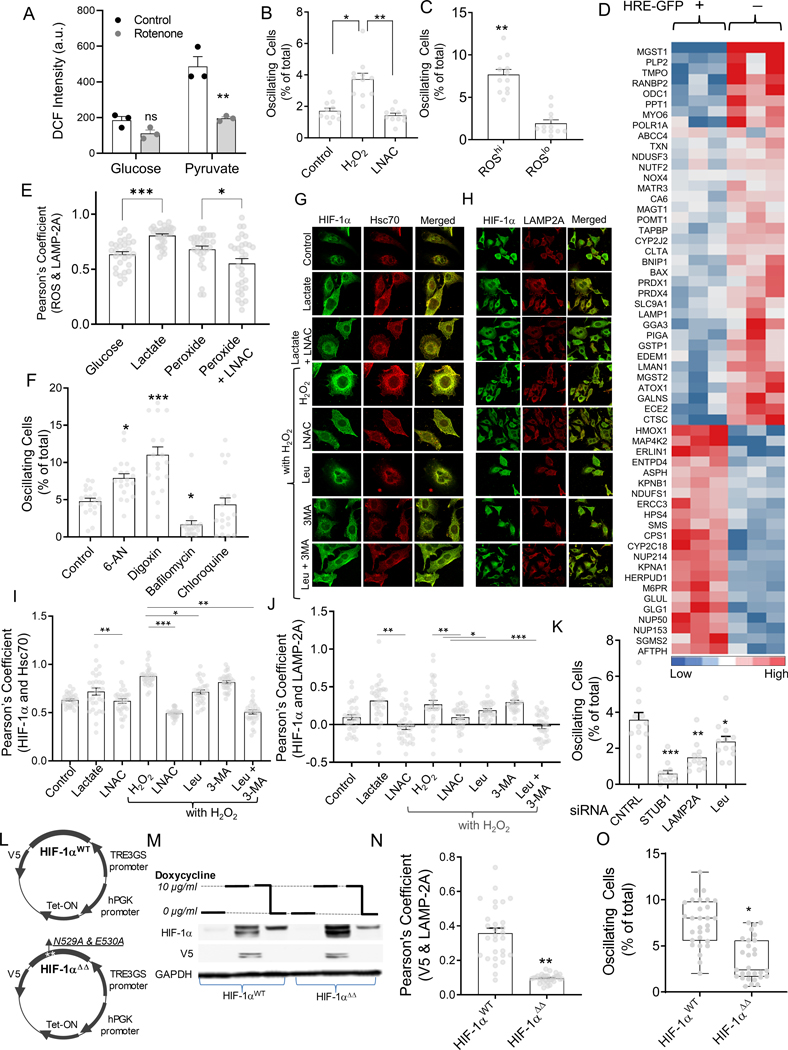
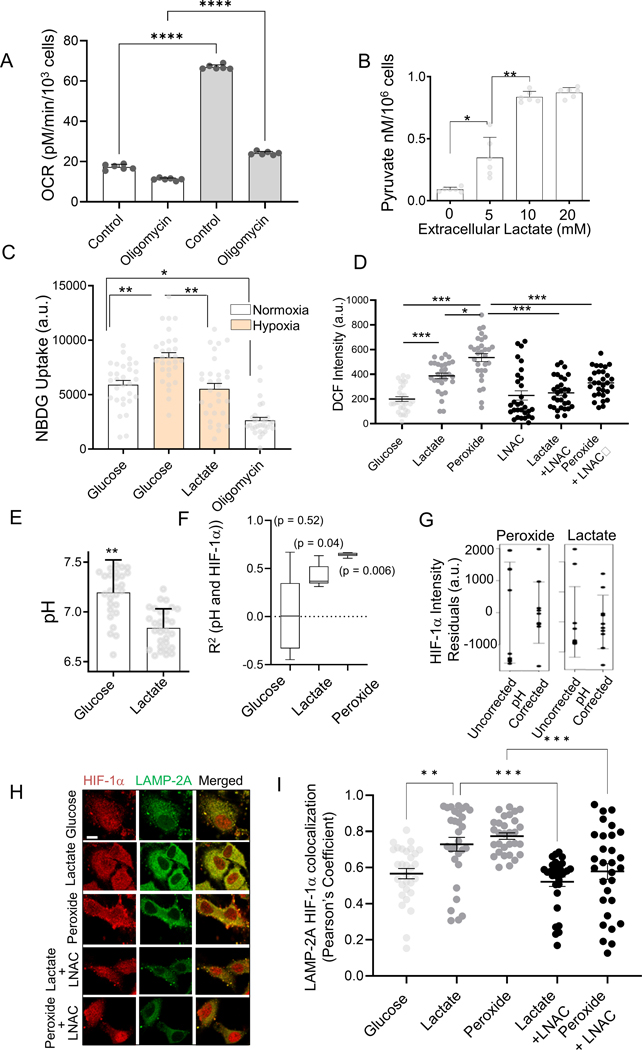
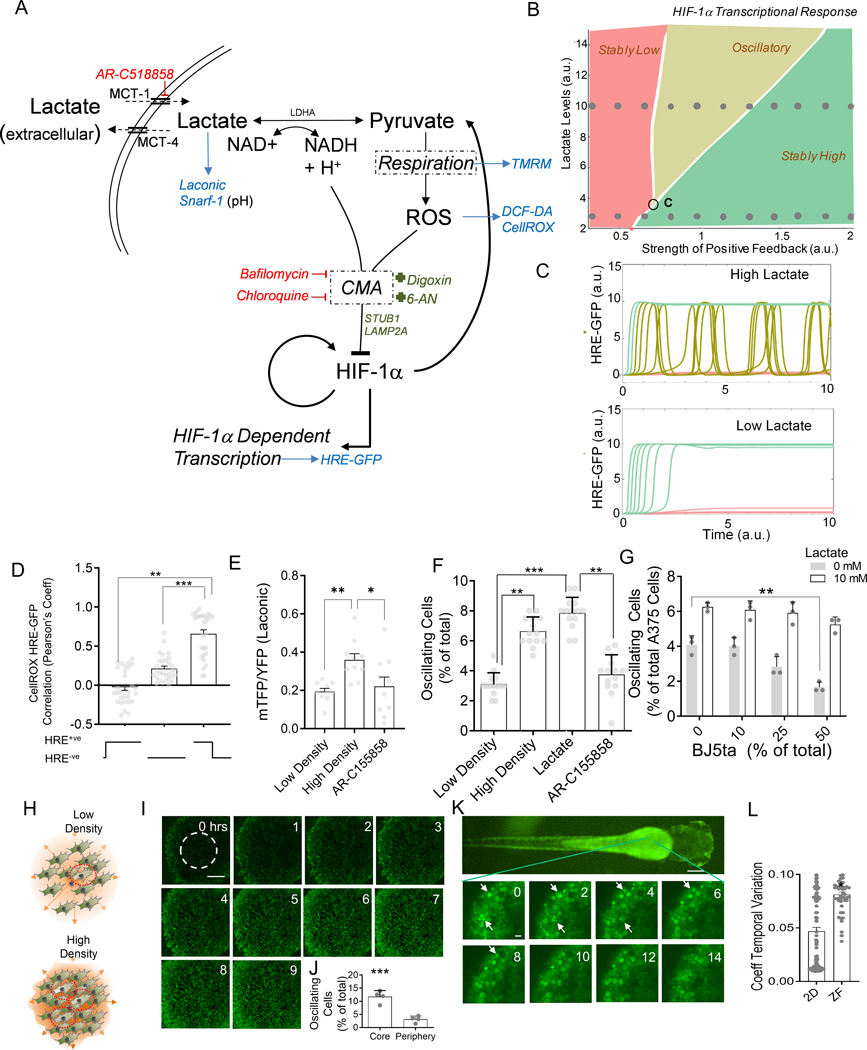
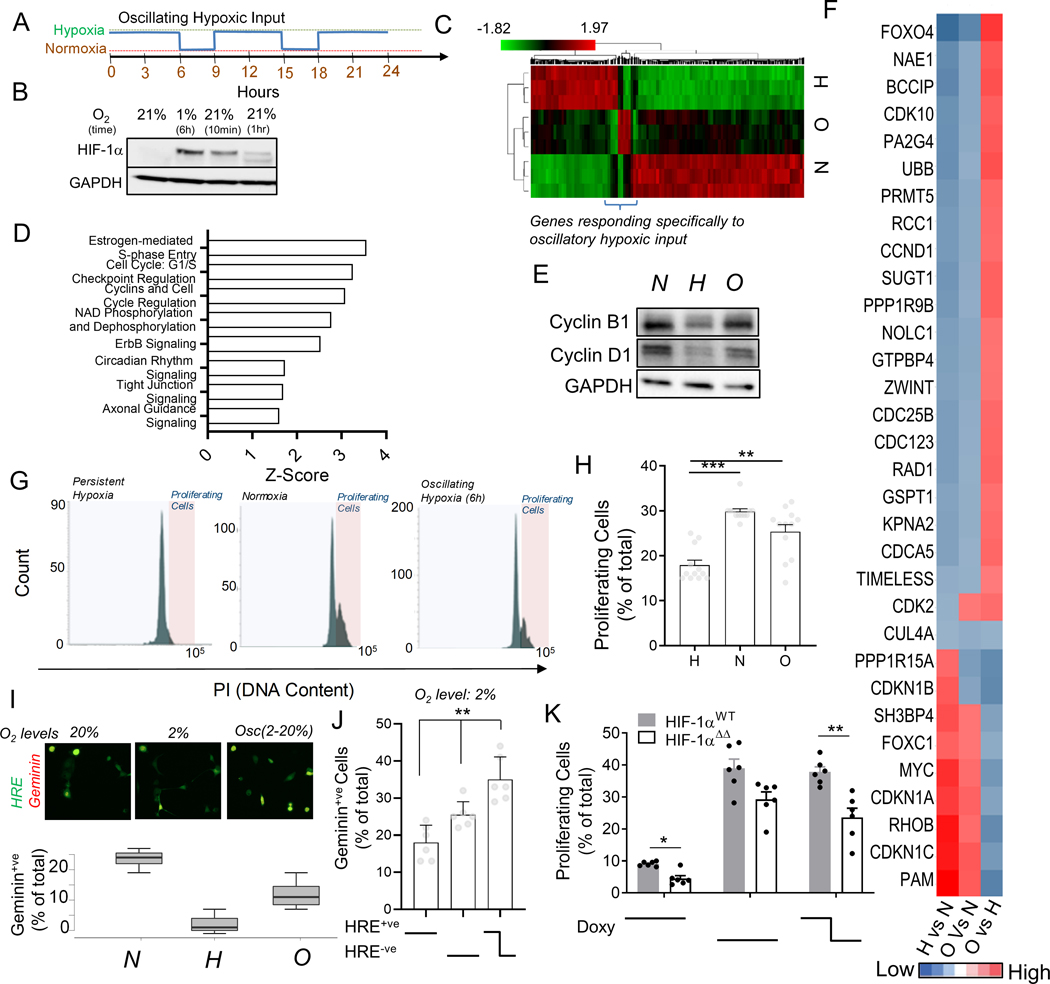
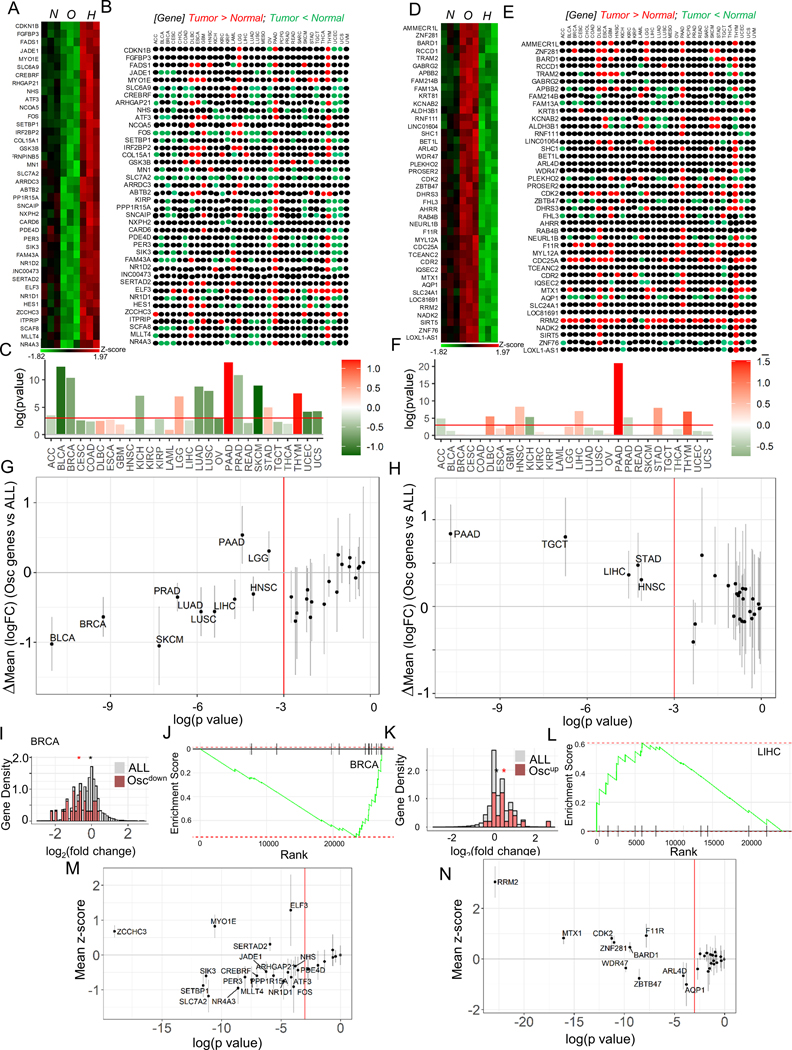
Response to hypoxia is a highly regulated process, but little is known about single-cell responses to hypoxic conditions. Using fluorescent reporters of hypoxia response factor-1α (HIF-1α) activity in various cancer cell lines and patient-derived cancer cells, we show that hypoxic responses in individual cancer cells can be highly dynamic and variable. These responses fall into three classes, including oscillatory activity. We identify a molecular mechanism that can account for all three response classes, implicating reactive-oxygen-species-dependent chaperone-mediated autophagy of HIF-1α in a subset of cells. Furthermore, we show that oscillatory response is modulated by the abundance of extracellular lactate in a quorum-sensing-like mechanism. We show that oscillatory HIF-1α activity rescues hypoxia-mediated inhibition of cell division and causes broad suppression of genes downregulated in cancers and activation of genes upregulated in many cancers, suggesting a mechanism for aggressive growth in a subset of hypoxic tumor cells.
Keywords: HIF; ROS; Warburg and reverse Warburg effect; cancer microenvironment; chaperone-mediated autophagy; hypoxia; hypoxic oscillations; lactate; quorum sensing; reactive oxygen species.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
