

Monocytes educated by cancer-associated fibroblasts secrete exosomal miR-181a to activate AKT signaling in breast cancer cells
- PMID: 36463188
- PMCID: PMC9719191
- DOI: 10.1186/s12967-022-03780-2
Monocytes educated by cancer-associated fibroblasts secrete exosomal miR-181a to activate AKT signaling in breast cancer cells
Abstract
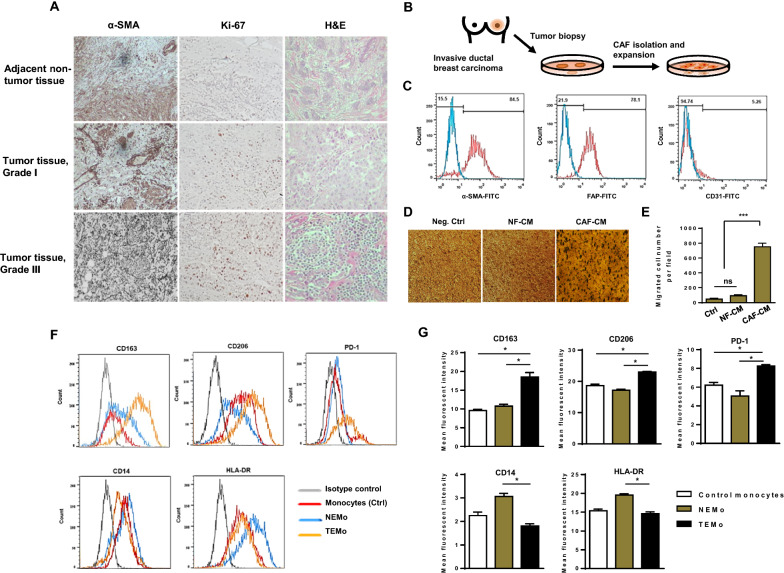
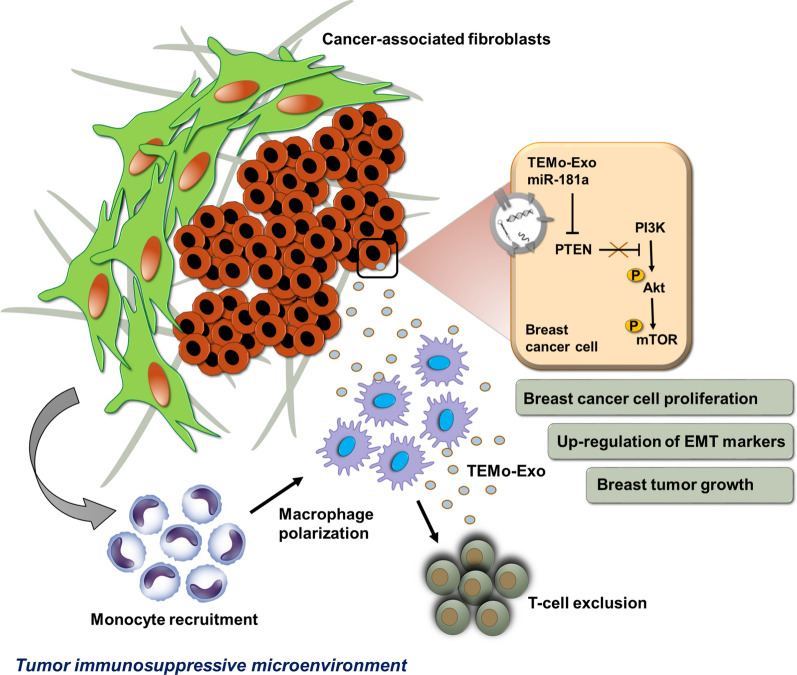
Background: Cancer-associated fibroblasts (CAFs), one of the major components of the tumor stroma, contribute to an immunosuppressive tumor microenvironment (TME) through the induction and functional polarization of protumoral macrophages. We have herein investigated the contribution of CAFs to monocyte recruitment and macrophage polarization. We also sought to identify a possible paracrine mechanism by which CAF-educated monocytes affect breast cancer (BC) cell progression.
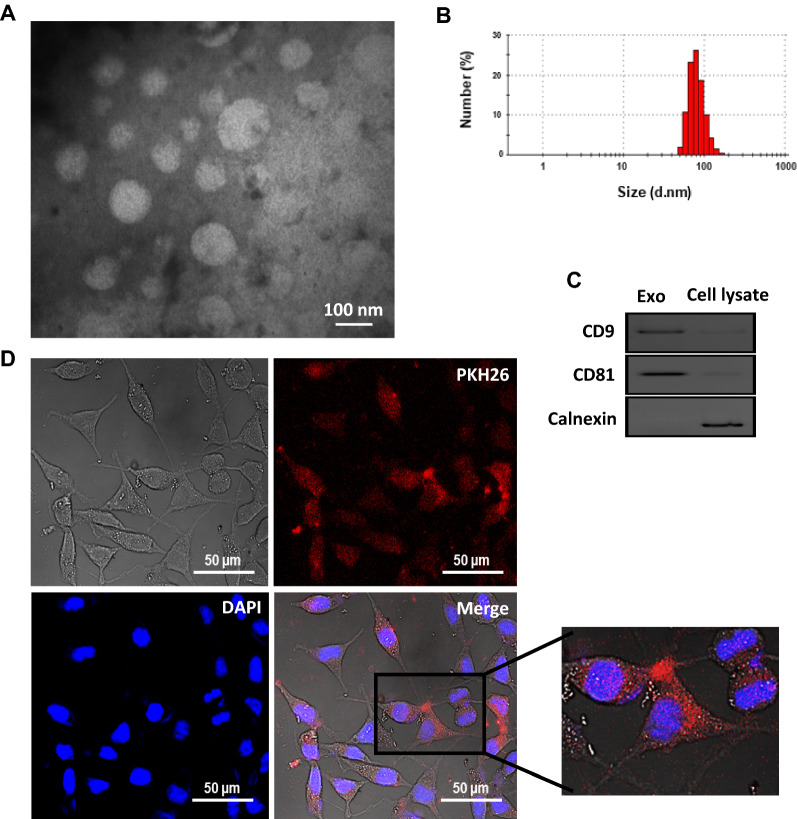
Methods: Monocytes were educated by primary CAFs and normal fibroblast (NF); the phenotypic alterations of CAF- or NF-educated monocytes were measured by flow cytometry. Exosomes isolated from the cultured conditioned media of the educated monocytes were characterized. An in vivo experiment using a subcutaneous transplantation tumor model in athymic nude mice was conducted to uncover the effect of exosomes derived from CAF- or NF-educated monocytes on breast tumor growth. Gain- and loss-of-function experiments were performed to explore the role of miR-181a in BC progression with the involvement of the AKT signaling pathway. Western blotting, enzyme-linked immunosorbent assay, RT-qPCR, flow cytometry staining, migration assay, immunohistochemical staining, and bioinformatics analysis were performed to reveal the underlying mechanisms.
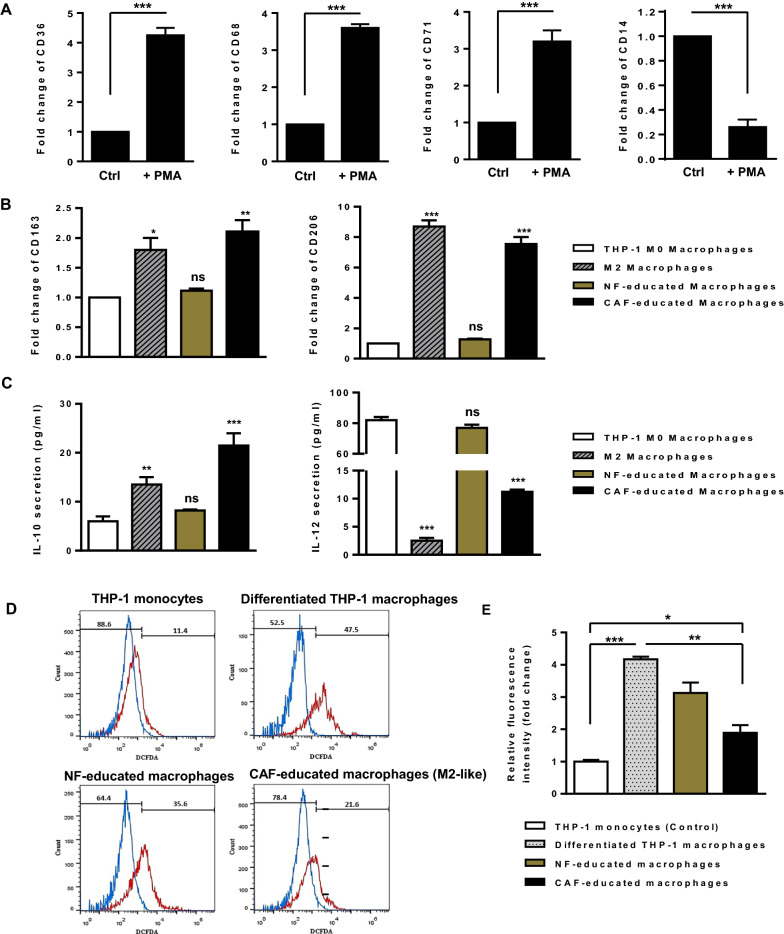
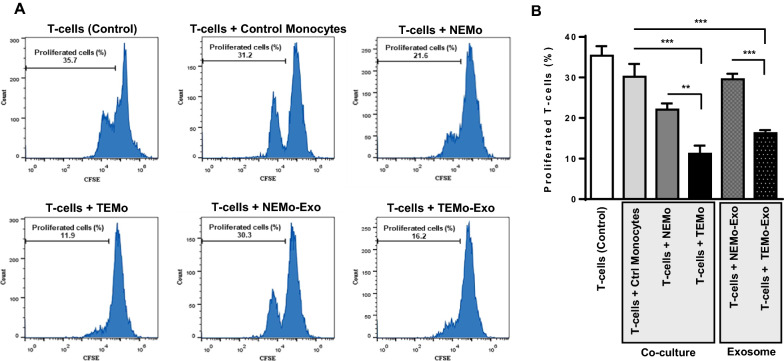
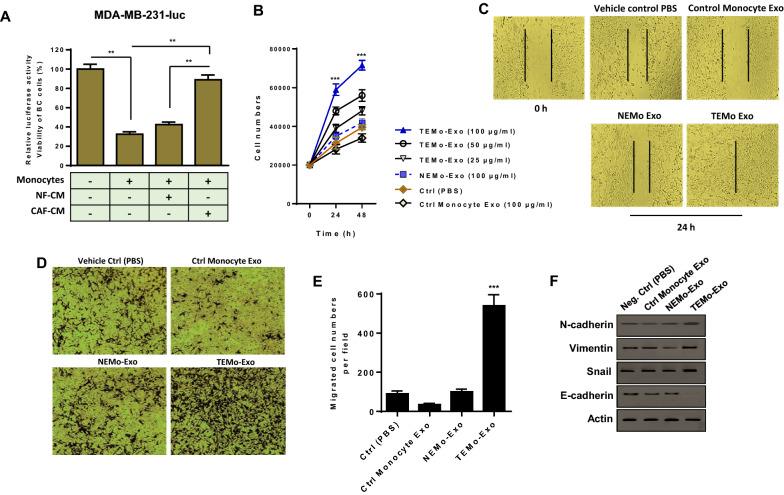
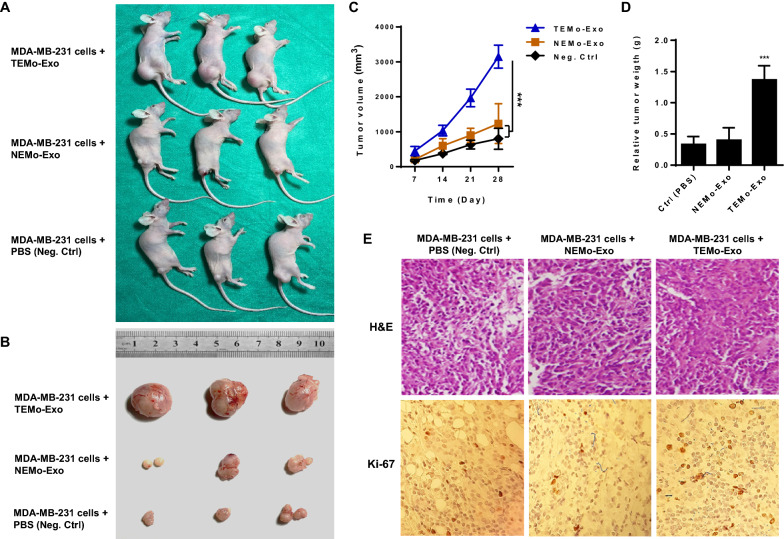
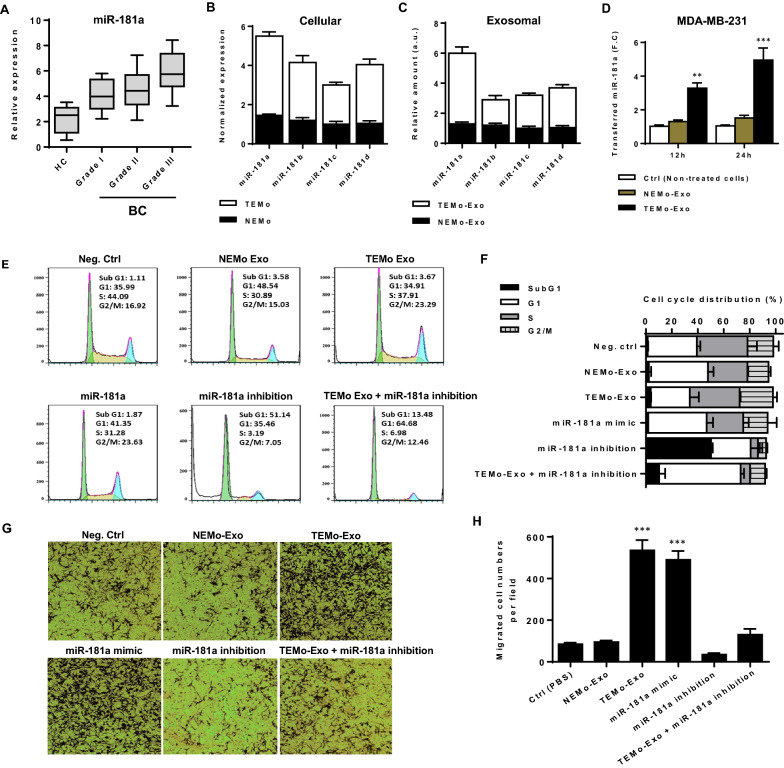
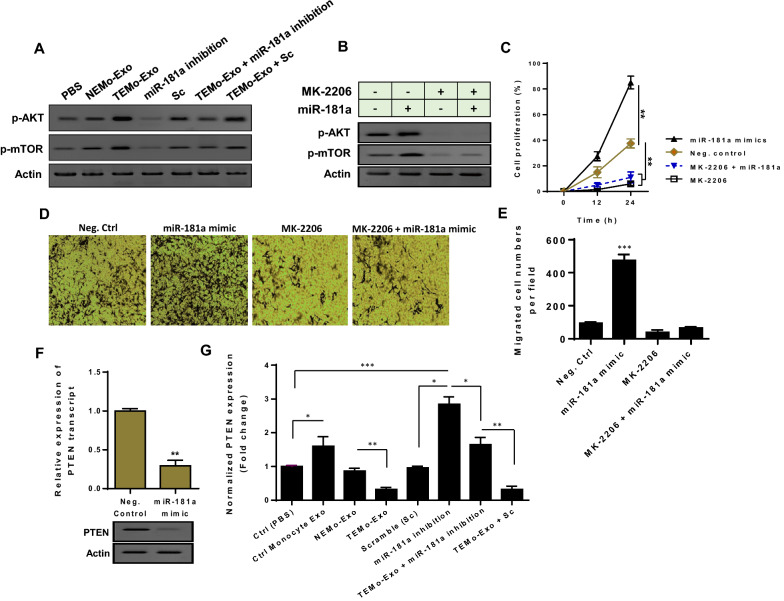
Results: We illustrated that primary CAFs recruited monocytes and established pro-tumoral M2 macrophages. CAF may also differentiate human monocyte THP-1 cells into anti-inflammatory M2 macrophages. Besides, we revealed that CAFs increased reactive oxygen species (ROS) generation in THP-1 monocytes, as differentiating into M2 macrophages requires a level of ROS for proper polarization. Importantly, T-cell proliferation was suppressed by CAF-educated monocytes and their exosomes, resulting in an immunosuppressive TME. Interestingly, CAF-activated, polarized monocytes lost their tumoricidal abilities, and their derived exosomes promoted BC cell proliferation and migration. In turn, CAF-educated monocyte exosomes exhibited a significant promoting effect on BC tumorigenicity in vivo. Of clinical significance, we observed that up-regulation of circulating miR-181a in BC was positively correlated with tumor aggressiveness and found a high level of this miRNA in CAF-educated monocytes and their exosomes. We further clarified that the pro-oncogenic effect of CAF-educated monocytes may depend in part on the exosomal transfer of miR-181a through modulating the PTEN/Akt signaling axis in BC cells.
Conclusions: Our findings established a connection between tumor stromal communication and tumor progression and demonstrated an inductive function for CAF-educated monocytes in BC cell progression. We also proposed a supporting model in which exosomal transfer of miR-181a from CAF-educated monocytes activates AKT signaling by regulating PTEN in BC cells.
Keywords: AKT signaling; Breast cancer; Cancer-associated fibroblasts; Exosomes; Immunosuppressive tumor microenvironment; Tumor-associated macrophages.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Cancer associated fibroblasts sculpt tumour microenvironment by recruiting monocytes and inducing immunosuppressive PD-1+ TAMs.Sci Rep. 2019 Feb 28;9(1):3172. doi: 10.1038/s41598-019-39553-z. Sci Rep. 2019. PMID: 30816272 Free PMC article.
-
CAFs-derived Exosomal miR-889-3p Might Repress M1 Macrophage Polarization to Boost ESCC Development by Regulating STAT1.Cell Biochem Biophys. 2025 Mar;83(1):633-646. doi: 10.1007/s12013-024-01496-2. Epub 2024 Sep 5. Cell Biochem Biophys. 2025. PMID: 39237779
-
Cancer-associated fibroblasts derived exosomal LINC01833 promotes the occurrence of non-small cell lung cancer through miR-335-5p -VAPA axis.J Biochem Mol Toxicol. 2024 Sep;38(9):e23769. doi: 10.1002/jbt.23769. J Biochem Mol Toxicol. 2024. PMID: 39152098
-
The role of CAF derived exosomal microRNAs in the tumour microenvironment of melanoma.Biochim Biophys Acta Rev Cancer. 2021 Jan;1875(1):188456. doi: 10.1016/j.bbcan.2020.188456. Epub 2020 Oct 22. Biochim Biophys Acta Rev Cancer. 2021. PMID: 33153973 Review.
-
Exosomal miRNAs and miRNA dysregulation in cancer-associated fibroblasts.Mol Cancer. 2017 Aug 29;16(1):148. doi: 10.1186/s12943-017-0718-4. Mol Cancer. 2017. PMID: 28851377 Free PMC article. Review.
Cited by
-
Exosomal miR-361-3p promotes the viability of breast cancer cells by targeting ETV7 and BATF2 to upregulate the PAI-1/ERK pathway.J Transl Med. 2024 Jan 28;22(1):112. doi: 10.1186/s12967-024-04914-4. J Transl Med. 2024. PMID: 38282047 Free PMC article.
-
Cancer-associated fibroblasts-derived exosomal piR-35462 promotes the progression of oral squamous cell carcinoma via FTO/Twist1 pathway.BMC Oral Health. 2025 May 28;25(1):840. doi: 10.1186/s12903-025-06082-3. BMC Oral Health. 2025. PMID: 40437442 Free PMC article.
-
Cancer-associated fibroblasts affect breast cancer cell sensitivity to chemotherapeutic agents by regulating NRBP2.Toxicol Res (Camb). 2024 Dec 8;13(6):tfae204. doi: 10.1093/toxres/tfae204. eCollection 2024 Dec. Toxicol Res (Camb). 2024. PMID: 39664500
-
Paracrine signaling in cancer-associated fibroblasts: central regulators of the tumor immune microenvironment.J Transl Med. 2025 Jun 23;23(1):697. doi: 10.1186/s12967-025-06744-4. J Transl Med. 2025. PMID: 40551209 Free PMC article. Review.
-
Extracellular vesicle-encapsulated microRNA-296-3p from cancer-associated fibroblasts promotes ovarian cancer development through regulation of the PTEN/AKT and SOCS6/STAT3 pathways.Cancer Sci. 2024 Jan;115(1):155-169. doi: 10.1111/cas.16014. Epub 2023 Nov 16. Cancer Sci. 2024. PMID: 37972389 Free PMC article.
References
-
- Costa A, Kieffer Y, Scholer-Dahirel A, Pelon F, Bourachot B, Cardon M, Sirven P, Magagna I, Fuhrmann L, Bernard C. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell. 2018;33(463–479):e410. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
