

Blockage of lamin-A/C loss diminishes the pro-inflammatory macrophage response
- PMID: 36465100
- PMCID: PMC9708799
- DOI: 10.1016/j.isci.2022.105528
Blockage of lamin-A/C loss diminishes the pro-inflammatory macrophage response
Abstract
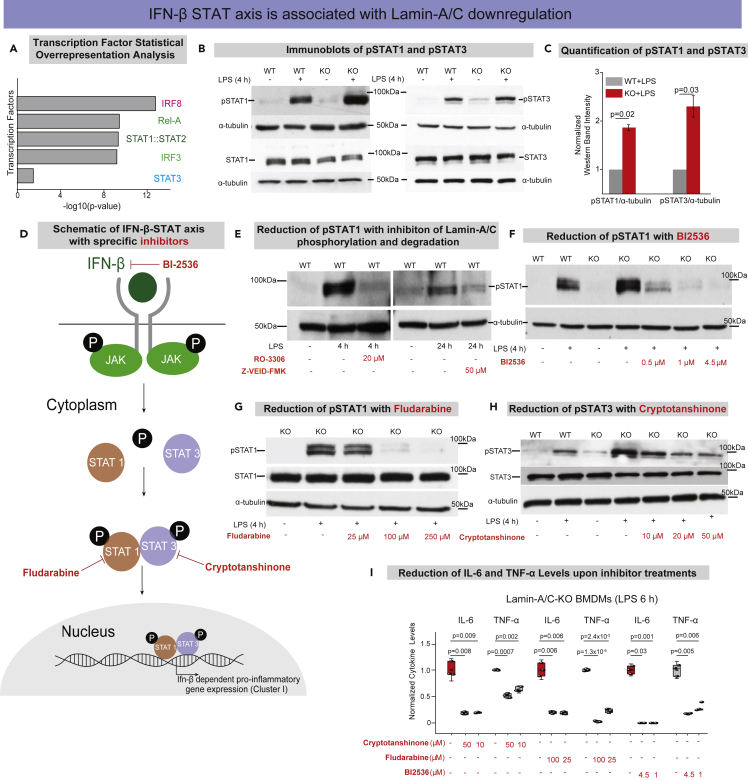
Mutations and defects in nuclear lamins can cause major pathologies, including inflammation and inflammatory diseases. Yet, the underlying molecular mechanisms are not known. We now report that the pro-inflammatory activation of macrophages, as induced by LPS or pathogenic E. coli, reduces Lamin-A/C levels thereby augmenting pro-inflammatory gene expression and cytokine secretion. We show that the activation of bone-marrow-derived macrophages (BMDMs) causes the phosphorylation and degradation of Lamin-A/C, as mediated by CDK1 and Caspase-6, respectively, necessary for upregulating IFN-β expression. Enhanced IFN-β expression subsequently increases pro-inflammatory gene expression via the IFN-β-STAT axis. Pro-inflammatory gene expression was also amplified in the complete absence of Lamin-A/C. Alternatively, pharmacological inhibition of either Lamin-A/C phosphorylation or degradation significantly downregulated pro-inflammatory gene expression, as did the targeting of IFN-β-STAT pathway members, i.e. phospho-STAT1 and phospho-STAT3. As Lamin-A/C is a previously unappreciated regulator of the pro-inflammatory macrophage response, our findings suggest novel opportunities to treat inflammatory diseases.
Keywords: Cell biology; Functional aspects of cell biology; Immune response; Immunology.
© 2022 The Author(s).
Conflict of interest statement
The authors declare that there is no conflict of interest regarding the publication of this article.
Figures





Similar articles
-
Lipocalin 2 is a regulator of macrophage polarization and NF-κB/STAT3 pathway activation.Mol Endocrinol. 2014 Oct;28(10):1616-28. doi: 10.1210/me.2014-1092. Epub 2014 Aug 15. Mol Endocrinol. 2014. PMID: 25127375 Free PMC article.
-
Luteolin transforms the polarity of bone marrow-derived macrophages to regulate the cytokine storm.J Inflamm (Lond). 2021 May 31;18(1):21. doi: 10.1186/s12950-021-00285-5. J Inflamm (Lond). 2021. PMID: 34059076 Free PMC article.
-
IFN-β Plays Both Pro- and Anti-inflammatory Roles in the Rat Cardiac Fibroblast Through Differential STAT Protein Activation.Front Pharmacol. 2018 Nov 28;9:1368. doi: 10.3389/fphar.2018.01368. eCollection 2018. Front Pharmacol. 2018. PMID: 30555324 Free PMC article.
-
STAT1 as a novel therapeutical target in pro-atherogenic signal integration of IFNγ, TLR4 and IL-6 in vascular disease.Cytokine Growth Factor Rev. 2011 Aug;22(4):211-9. doi: 10.1016/j.cytogfr.2011.06.003. Epub 2011 Jul 12. Cytokine Growth Factor Rev. 2011. PMID: 21752694 Review.
-
Nuclear lamin phosphorylation: an emerging role in gene regulation and pathogenesis of laminopathies.Nucleus. 2020 Dec;11(1):299-314. doi: 10.1080/19491034.2020.1832734. Nucleus. 2020. PMID: 33030403 Free PMC article. Review.
Cited by
-
Intermediate, but not average: The unusual lives of the nuclear lamin proteins.Curr Opin Cell Biol. 2023 Oct;84:102220. doi: 10.1016/j.ceb.2023.102220. Epub 2023 Aug 22. Curr Opin Cell Biol. 2023. PMID: 37619289 Free PMC article. Review.
-
Clustered macrophages cooperate to eliminate tumors via coordinated intrudopodia.Proc Natl Acad Sci U S A. 2025 Jul 8;122(27):e2425452122. doi: 10.1073/pnas.2425452122. Epub 2025 Jul 1. Proc Natl Acad Sci U S A. 2025. PMID: 40591598
-
Regulation of apoptosis and interaction with cartilage degeneration in osteoarthritis.Front Cell Dev Biol. 2025 Mar 27;13:1571448. doi: 10.3389/fcell.2025.1571448. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40213395 Free PMC article. Review.
-
Targeting Nuclear Mechanics Mitigates the Fibroblast Invasiveness in Pathological Dermal Scars Induced by Matrix Stiffening.Adv Sci (Weinh). 2024 Apr;11(15):e2308253. doi: 10.1002/advs.202308253. Epub 2024 Feb 14. Adv Sci (Weinh). 2024. PMID: 38353381 Free PMC article.
-
Mechanoimmunology: Are inflammatory epigenetic states of macrophages tuned by biophysical factors?APL Bioeng. 2022 Aug 29;6(3):031502. doi: 10.1063/5.0087699. eCollection 2022 Sep. APL Bioeng. 2022. PMID: 36051106 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
