

S945L-CFTR molecular dynamics, functional characterization and tezacaftor/ivacaftor efficacy in vivo and in vitro in matched pediatric patient-derived cell models
- PMID: 36467478
- PMCID: PMC9709344
- DOI: 10.3389/fped.2022.1062766
S945L-CFTR molecular dynamics, functional characterization and tezacaftor/ivacaftor efficacy in vivo and in vitro in matched pediatric patient-derived cell models
Abstract
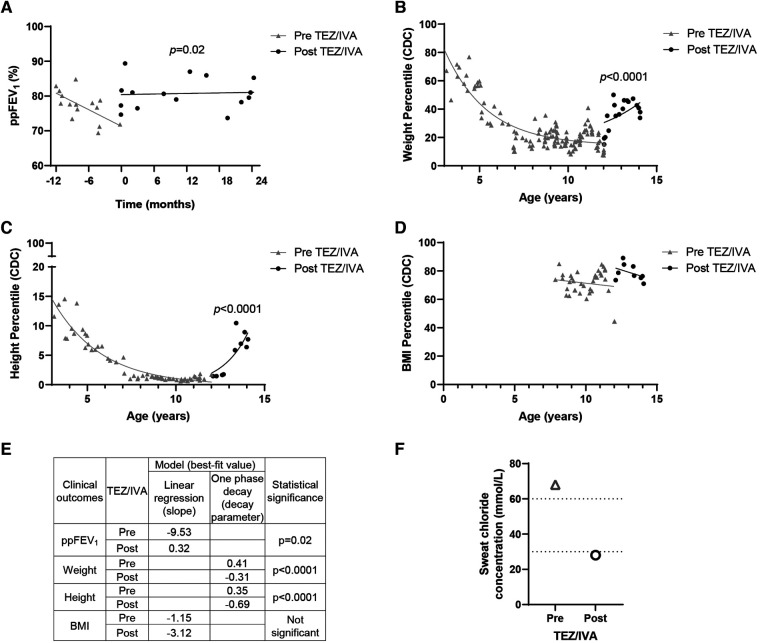
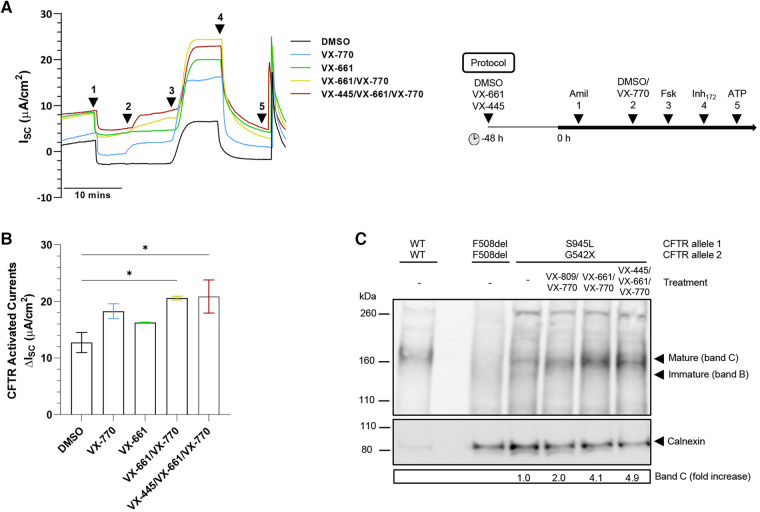
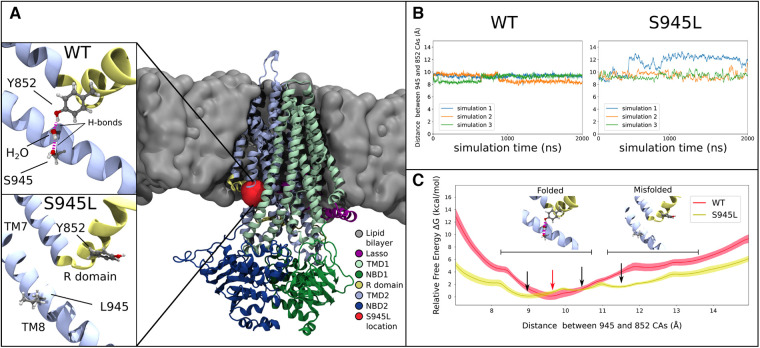
Cystic Fibrosis (CF) results from over 400 different disease-causing mutations in the CF Transmembrane Conductance Regulator (CFTR) gene. These CFTR mutations lead to numerous defects in CFTR protein function. A novel class of targeted therapies (CFTR modulators) have been developed that can restore defects in CFTR folding and gating. This study aimed to characterize the functional and structural defects of S945L-CFTR and interrogate the efficacy of modulators with two modes of action: gating potentiator [ivacaftor (IVA)] and folding corrector [tezacaftor (TEZ)]. The response to these modulators in vitro in airway differentiated cell models created from a participant with S945L/G542X-CFTR was correlated with in vivo clinical outcomes of that participant at least 12 months pre and post modulator therapy. In this participants' airway cell models, CFTR-mediated chloride transport was assessed via ion transport electrophysiology. Monotherapy with IVA or TEZ increased CFTR activity, albeit not reaching statistical significance. Combination therapy with TEZ/IVA significantly (p = 0.02) increased CFTR activity 1.62-fold above baseline. Assessment of CFTR expression and maturation via western blot validated the presence of mature, fully glycosylated CFTR, which increased 4.1-fold in TEZ/IVA-treated cells. The in vitro S945L-CFTR response to modulator correlated with an improvement in in vivo lung function (ppFEV1) from 77.19 in the 12 months pre TEZ/IVA to 80.79 in the 12 months post TEZ/IVA. The slope of decline in ppFEV1 significantly (p = 0.02) changed in the 24 months post TEZ/IVA, becoming positive. Furthermore, there was a significant improvement in clinical parameters and a fall in sweat chloride from 68 to 28 mmol/L. The mechanism of dysfunction of S945L-CFTR was elucidated by in silico molecular dynamics (MD) simulations. S945L-CFTR caused misfolding of transmembrane helix 8 and disruption of the R domain, a CFTR domain critical to channel gating. This study showed in vitro and in silico that S945L causes both folding and gating defects in CFTR and demonstrated in vitro and in vivo that TEZ/IVA is an efficacious modulator combination to address these defects. As such, we support the utility of patient-derived cell models and MD simulations in predicting and understanding the effect of modulators on CFTR function on an individualized basis.
Keywords: CFTR; airway epithelial cell models; cystic fibrosis; modulators; molecular dynamics; personalized medicine.
© 2022 Allan, Astore, Fawcett, Wong, Chen, Griffith, Jaffe, Kuyucak and Waters.
Conflict of interest statement
SAW is the recipient of a Vertex Innovation Grant (2018) and a TSANZ/Vertex Research Award (2020). Both are unrelated and outside of the submitted manuscript. AJ has received consulting fees from Vertex on projects unrelated to this study. All other authors declare no conflict of interest.
Figures
Similar articles
-
A phase 3, randomized, double-blind, parallel-group study to evaluate tezacaftor/ivacaftor in people with cystic fibrosis heterozygous for F508del-CFTR and a gating mutation.J Cyst Fibros. 2021 Mar;20(2):234-242. doi: 10.1016/j.jcf.2020.11.003. Epub 2020 Dec 16. J Cyst Fibros. 2021. PMID: 33339768 Clinical Trial.
-
Effects of Elexacaftor/Tezacaftor/Ivacaftor Therapy on CFTR Function in Patients with Cystic Fibrosis and One or Two F508del Alleles.Am J Respir Crit Care Med. 2022 Mar 1;205(5):540-549. doi: 10.1164/rccm.202110-2249OC. Am J Respir Crit Care Med. 2022. PMID: 34936849
-
Personalized medicine with drugs targeting the underlying protein defect in cystic fibrosis: is monitoring of treatment response necessary?Ther Adv Chronic Dis. 2022 Aug 5;13:20406223221108627. doi: 10.1177/20406223221108627. eCollection 2022. Ther Adv Chronic Dis. 2022. PMID: 35959505 Free PMC article.
-
Progress of personalized medicine of cystic fibrosis in the times of efficient CFTR modulators.Mol Cell Pediatr. 2025 May 5;12(1):6. doi: 10.1186/s40348-025-00194-0. Mol Cell Pediatr. 2025. PMID: 40320452 Free PMC article. Review.
-
Efficacy and Safety of Elexacaftor-Tezacaftor-Ivacaftor in the Treatment of Cystic Fibrosis: A Systematic Review.Children (Basel). 2023 Mar 15;10(3):554. doi: 10.3390/children10030554. Children (Basel). 2023. PMID: 36980112 Free PMC article. Review.
Cited by
-
Comparing Cytology Brushes for Optimal Human Nasal Epithelial Cell Collection: Implications for Airway Disease Diagnosis and Research.J Pers Med. 2023 May 21;13(5):864. doi: 10.3390/jpm13050864. J Pers Med. 2023. PMID: 37241034 Free PMC article.
-
Rescue of Mutant CFTR Channel Activity by Investigational Co-Potentiator Therapy.Biomedicines. 2025 Jan 1;13(1):82. doi: 10.3390/biomedicines13010082. Biomedicines. 2025. PMID: 39857666 Free PMC article.
-
Molecular and Functional Characteristics of Airway Epithelium under Chronic Hypoxia.Int J Mol Sci. 2023 Mar 30;24(7):6475. doi: 10.3390/ijms24076475. Int J Mol Sci. 2023. PMID: 37047450 Free PMC article.
-
Q1291H-CFTR molecular dynamics simulations and ex vivo theratyping in nasal epithelial models and clinical response to elexacaftor/tezacaftor/ivacaftor in a Q1291H/F508del patient.Front Mol Biosci. 2023 Jun 1;10:1148501. doi: 10.3389/fmolb.2023.1148501. eCollection 2023. Front Mol Biosci. 2023. PMID: 37325471 Free PMC article.
References
-
- https://www.cftr2.org/ The Clinical and Functional Translation of Cftr (Cftr2). US CF Foundation, Johns Hopkins University, and The Hospital for Sick Children (2011). Available at:
LinkOut - more resources
Full Text Sources
Research Materials