

Desiccation resistance differences in Drosophila species can be largely explained by variations in cuticular hydrocarbons
- PMID: 36473178
- PMCID: PMC9757832
- DOI: 10.7554/eLife.80859
Desiccation resistance differences in Drosophila species can be largely explained by variations in cuticular hydrocarbons
Abstract
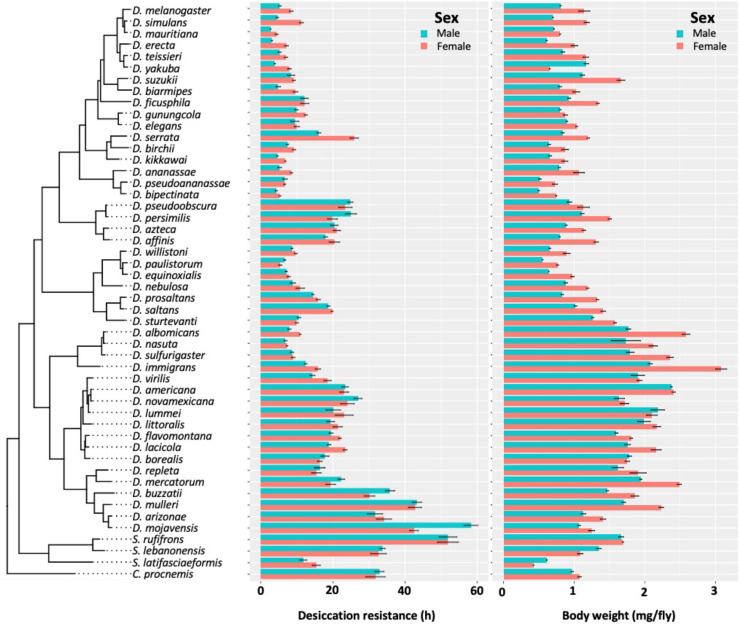
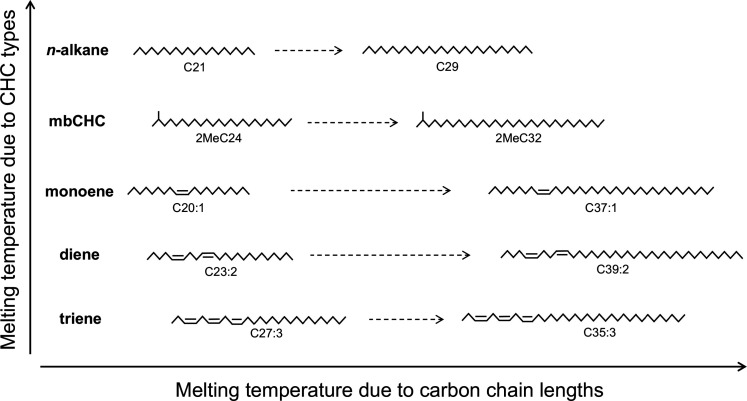
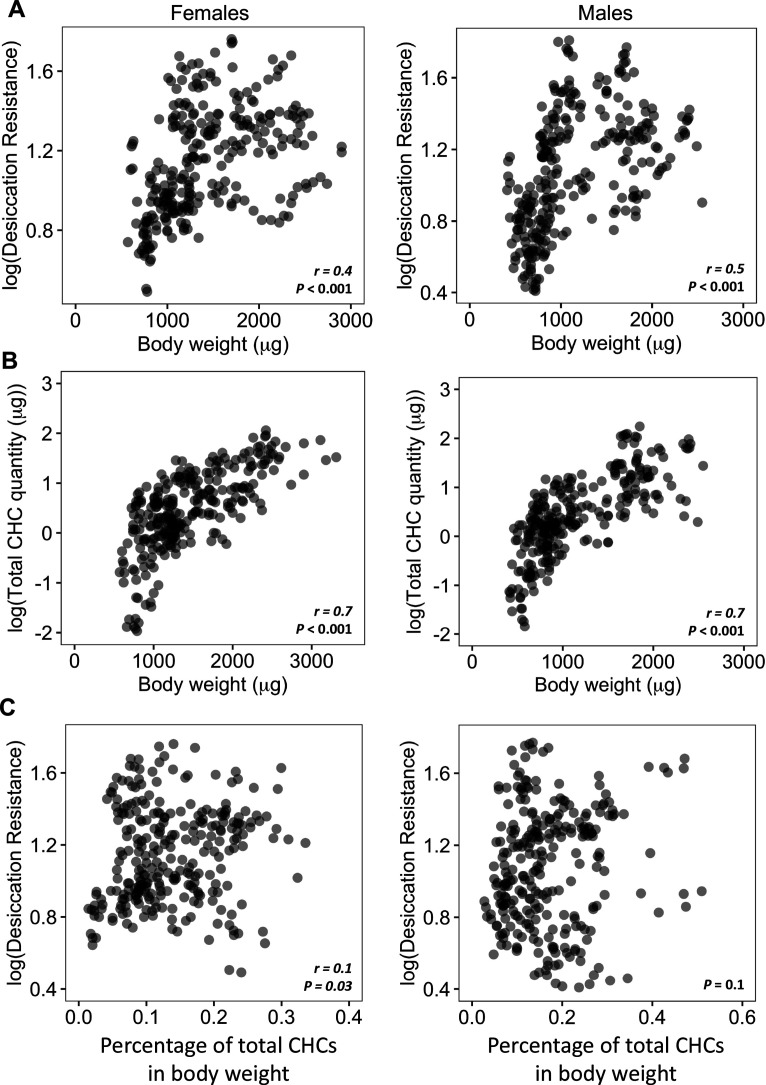
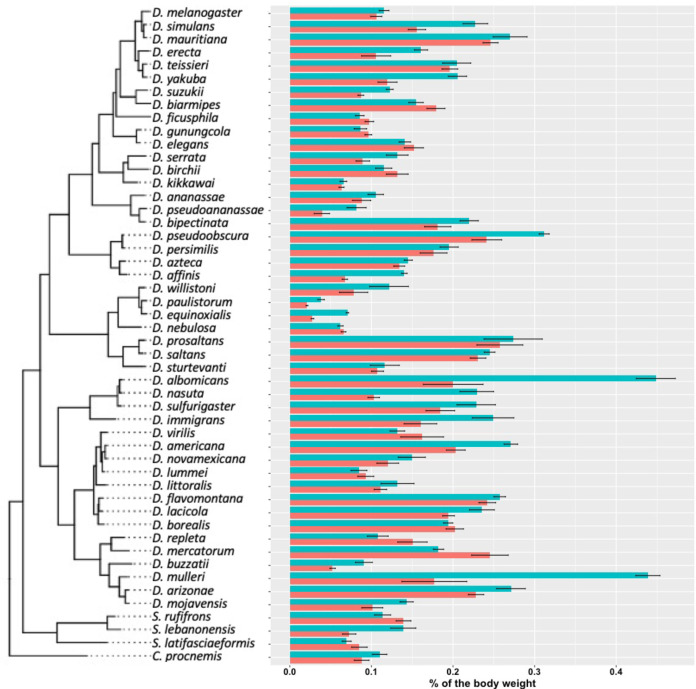
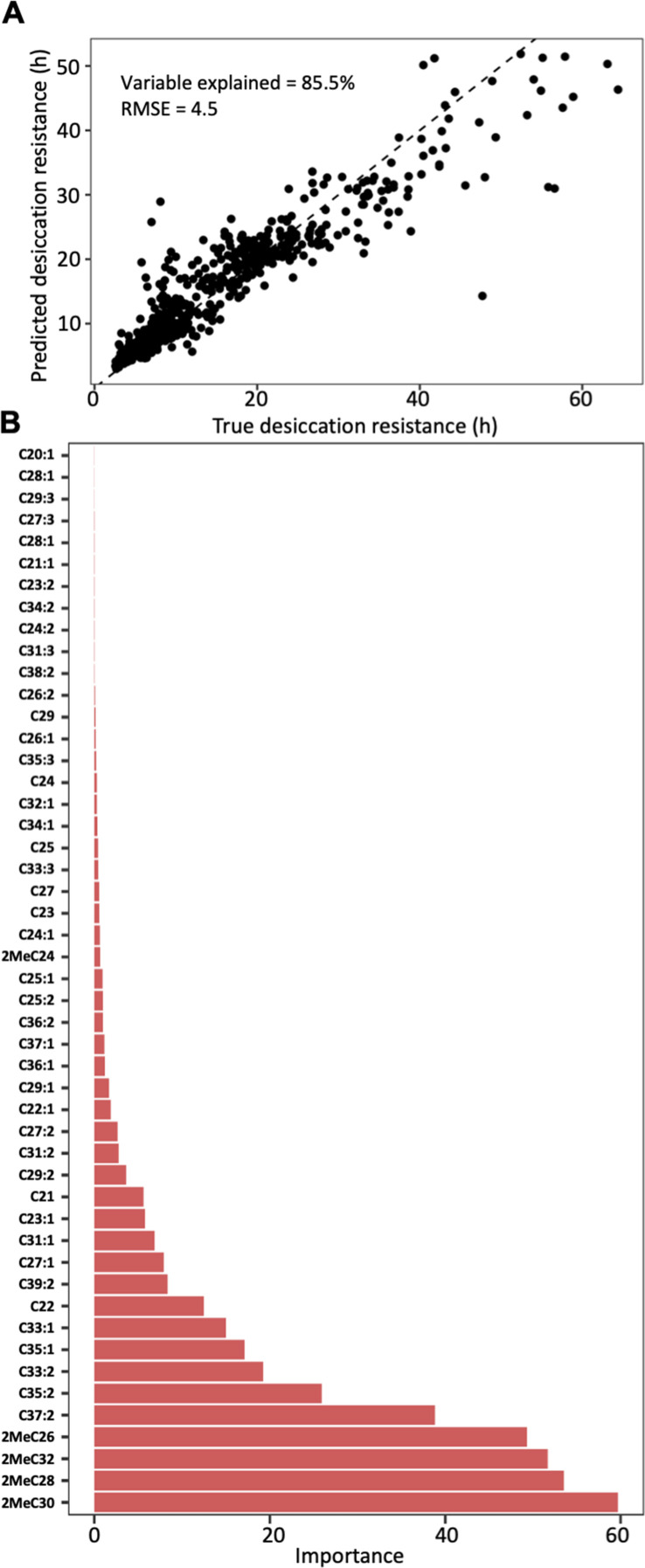
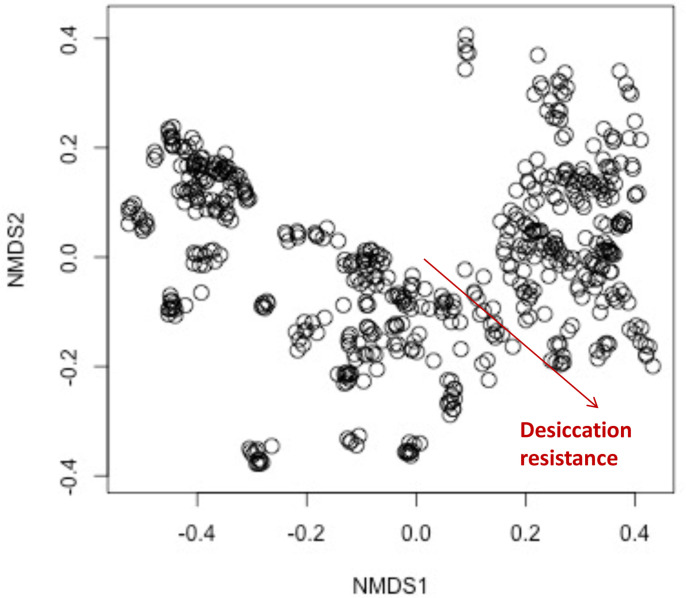
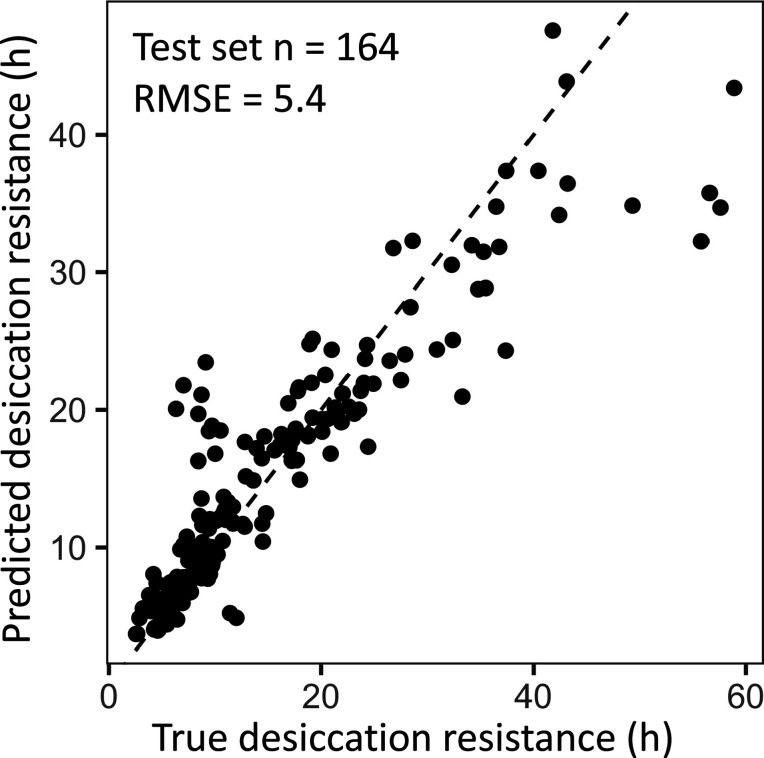
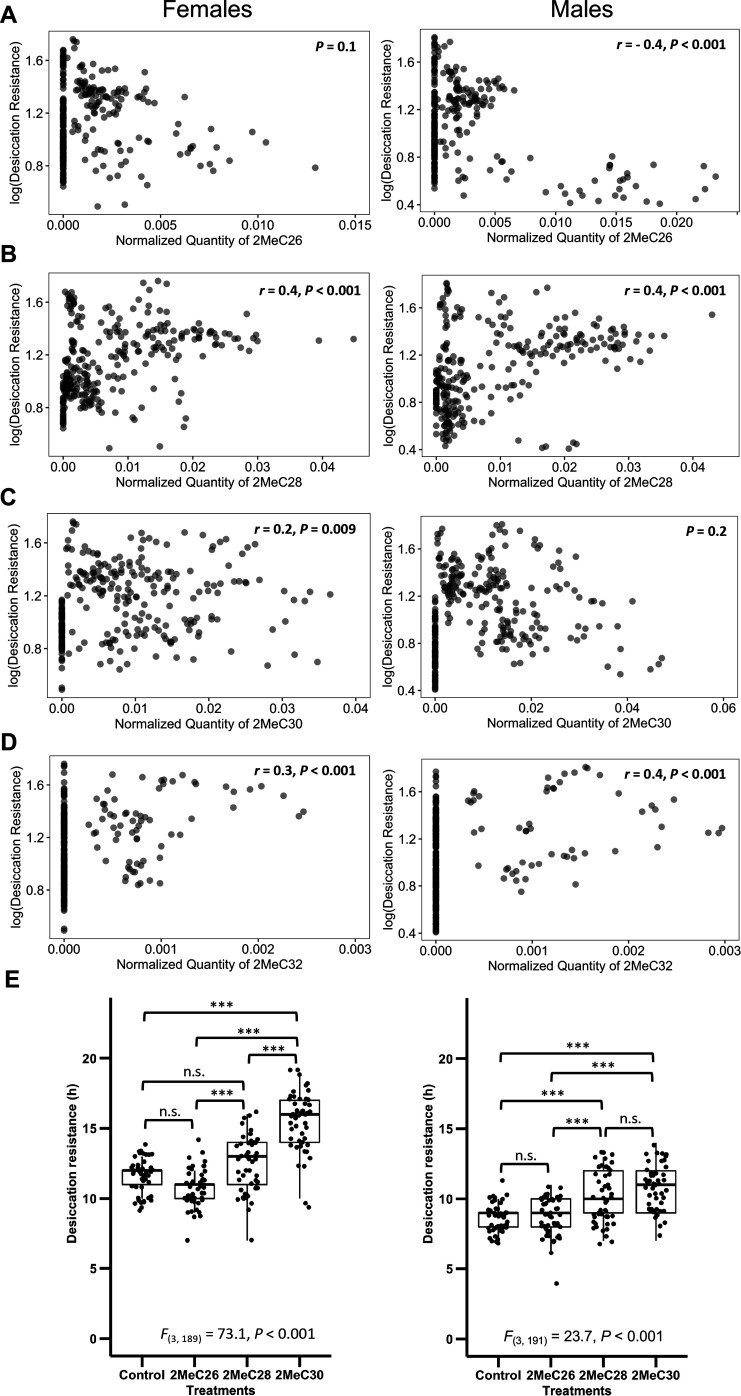
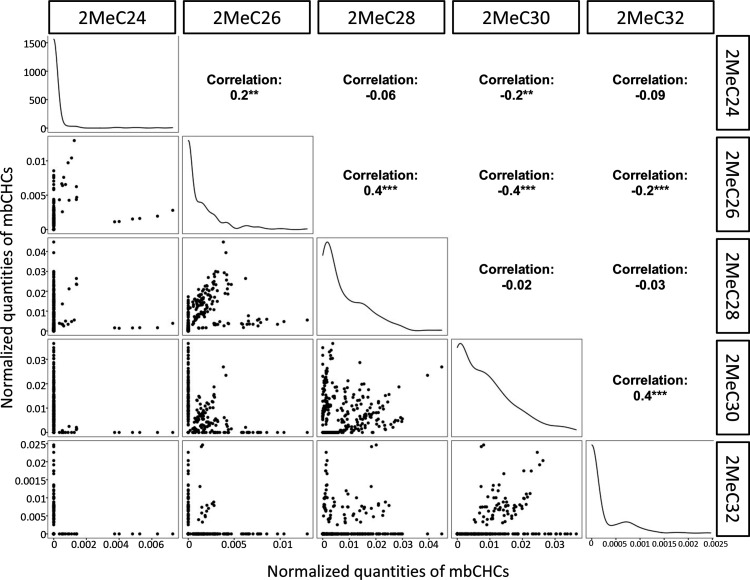
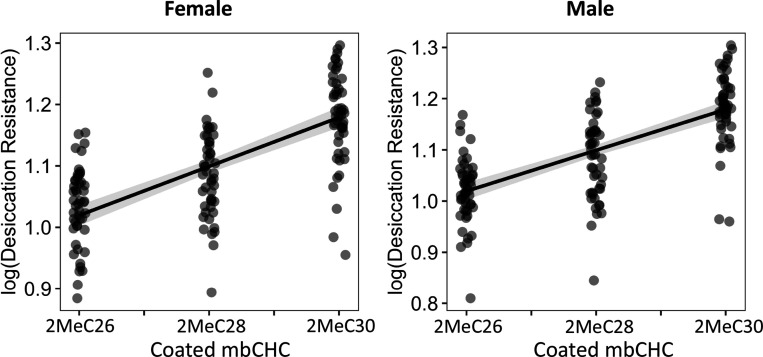
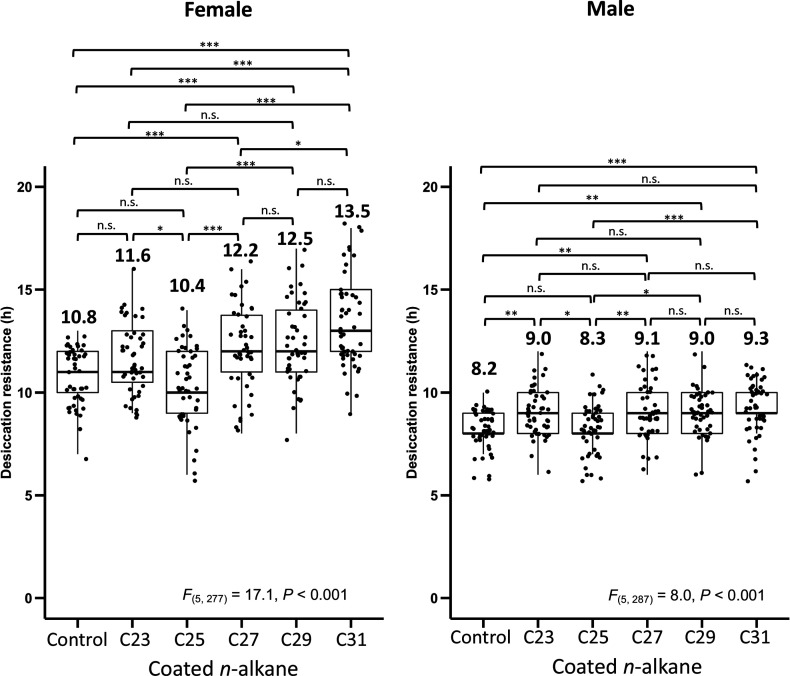
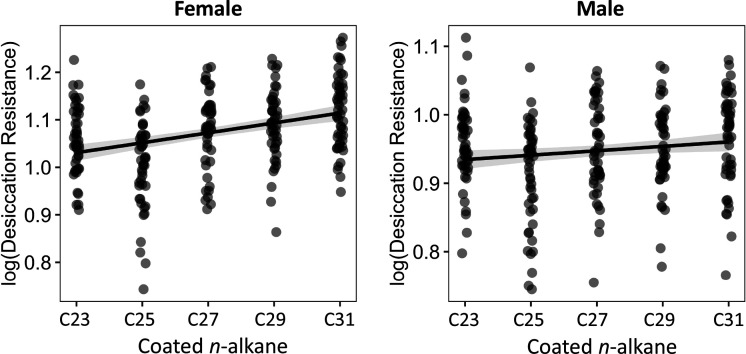
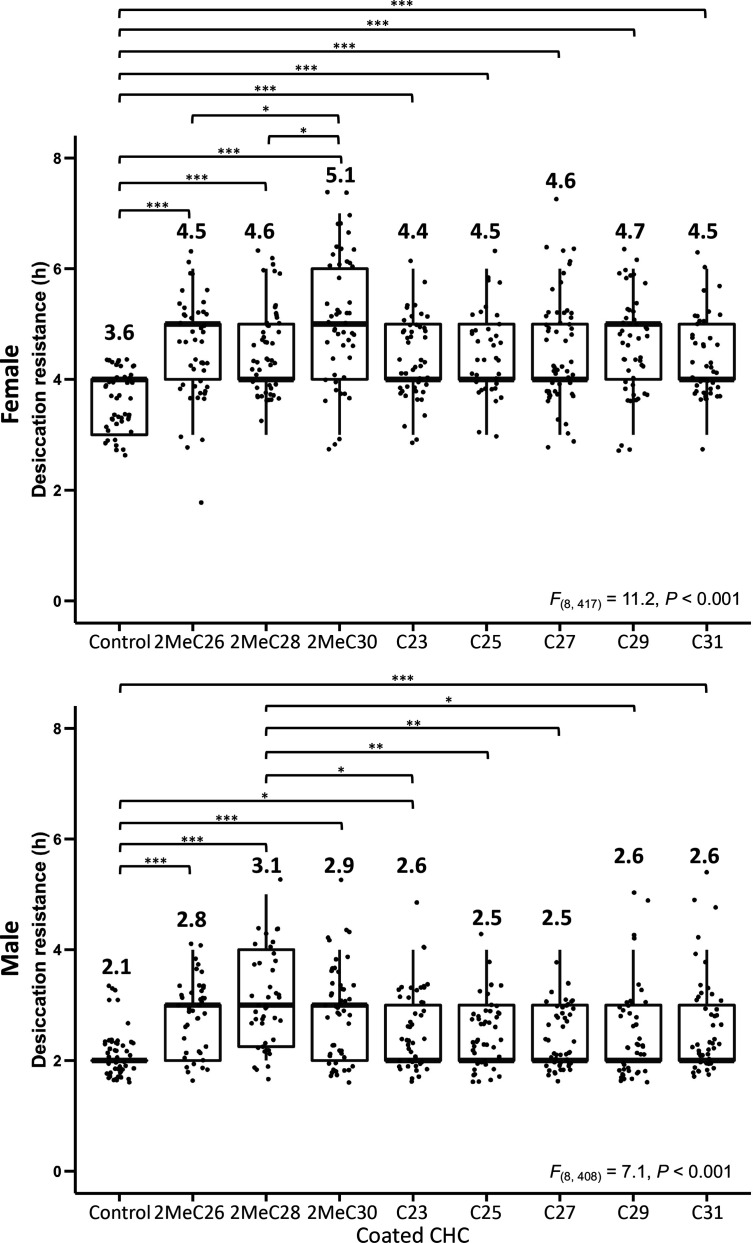
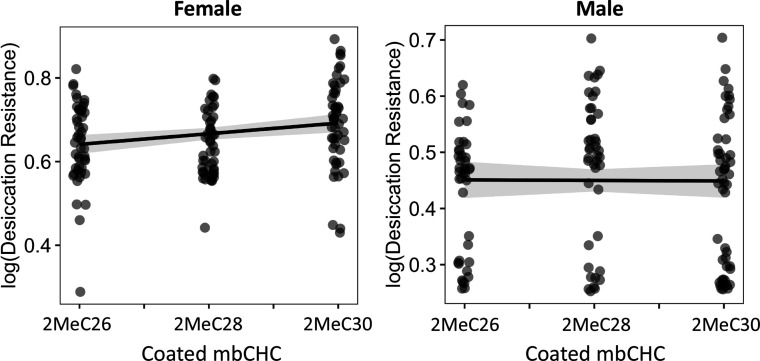
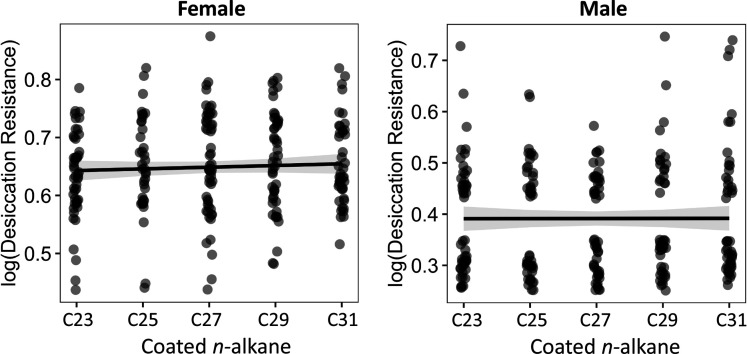
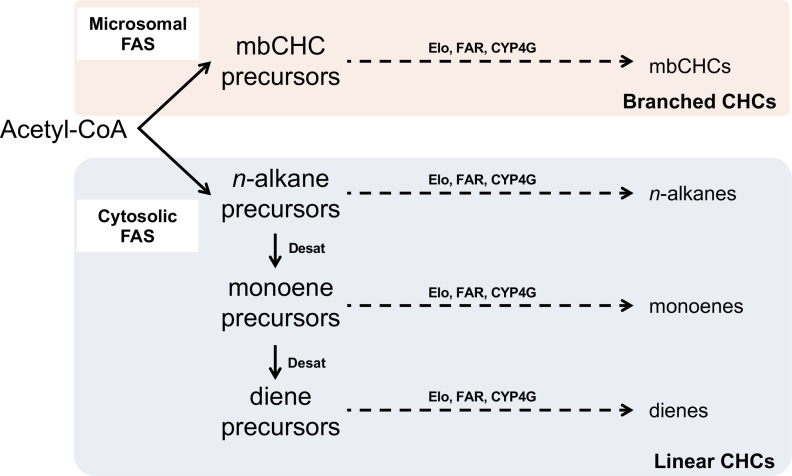
Maintaining water balance is a universal challenge for organisms living in terrestrial environments, especially for insects, which have essential roles in our ecosystem. Although the high surface area to volume ratio in insects makes them vulnerable to water loss, insects have evolved different levels of desiccation resistance to adapt to diverse environments. To withstand desiccation, insects use a lipid layer called cuticular hydrocarbons (CHCs) to reduce water evaporation from the body surface. It has long been hypothesized that the water-proofing capability of this CHC layer, which can confer different levels of desiccation resistance, depends on its chemical composition. However, it is unknown which CHC components are important contributors to desiccation resistance and how these components can determine differences in desiccation resistance. In this study, we used machine-learning algorithms, correlation analyses, and synthetic CHCs to investigate how different CHC components affect desiccation resistance in 50 Drosophila and related species. We showed that desiccation resistance differences across these species can be largely explained by variation in CHC composition. In particular, length variation in a subset of CHCs, the methyl-branched CHCs (mbCHCs), is a key determinant of desiccation resistance. There is also a significant correlation between the evolution of longer mbCHCs and higher desiccation resistance in these species. Given that CHCs are almost ubiquitous in insects, we suggest that evolutionary changes in insect CHC components can be a general mechanism for the evolution of desiccation resistance and adaptation to diverse and changing environments.
Keywords: D. melanogaster; Drosophila species; ecology; evolutionary biology; methyl-branched cuticular hydrocarbons; none; random forest.
© 2022, Wang et al.
Conflict of interest statement
ZW, JR, JP, HC, CR, ML, HC No competing interests declared
Figures
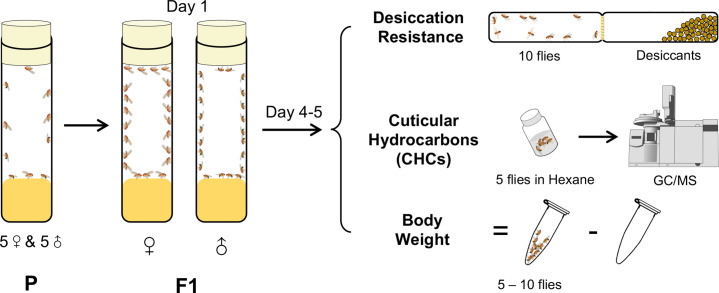
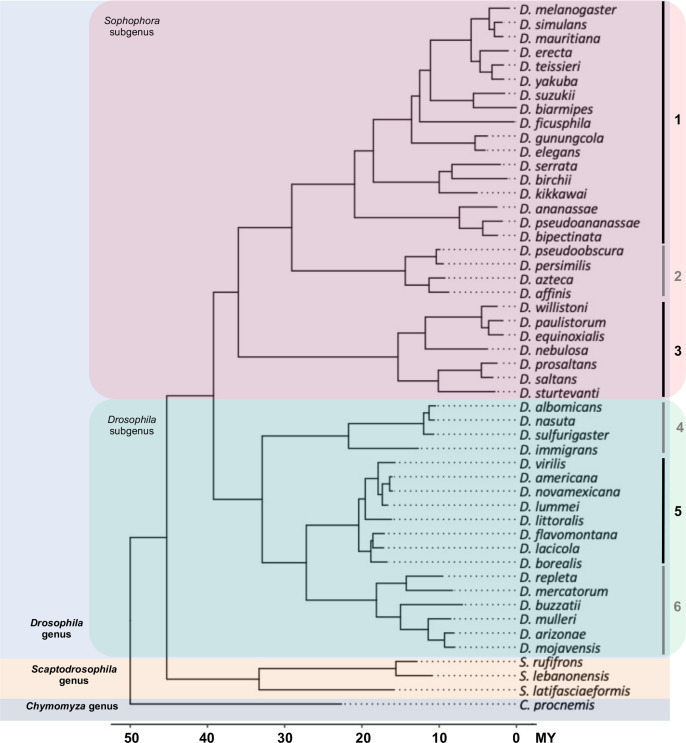
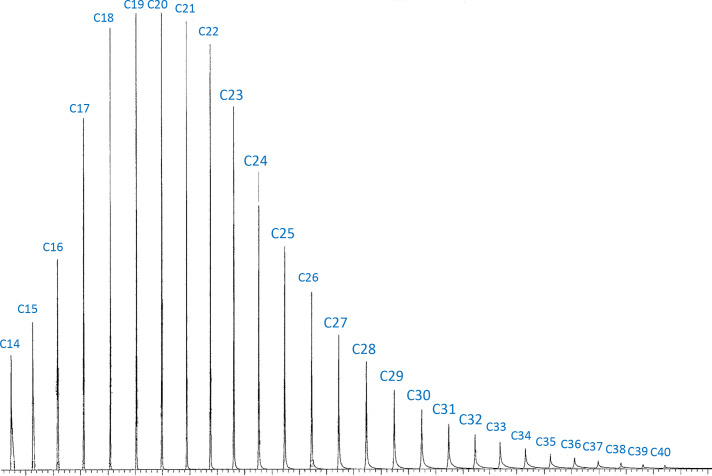
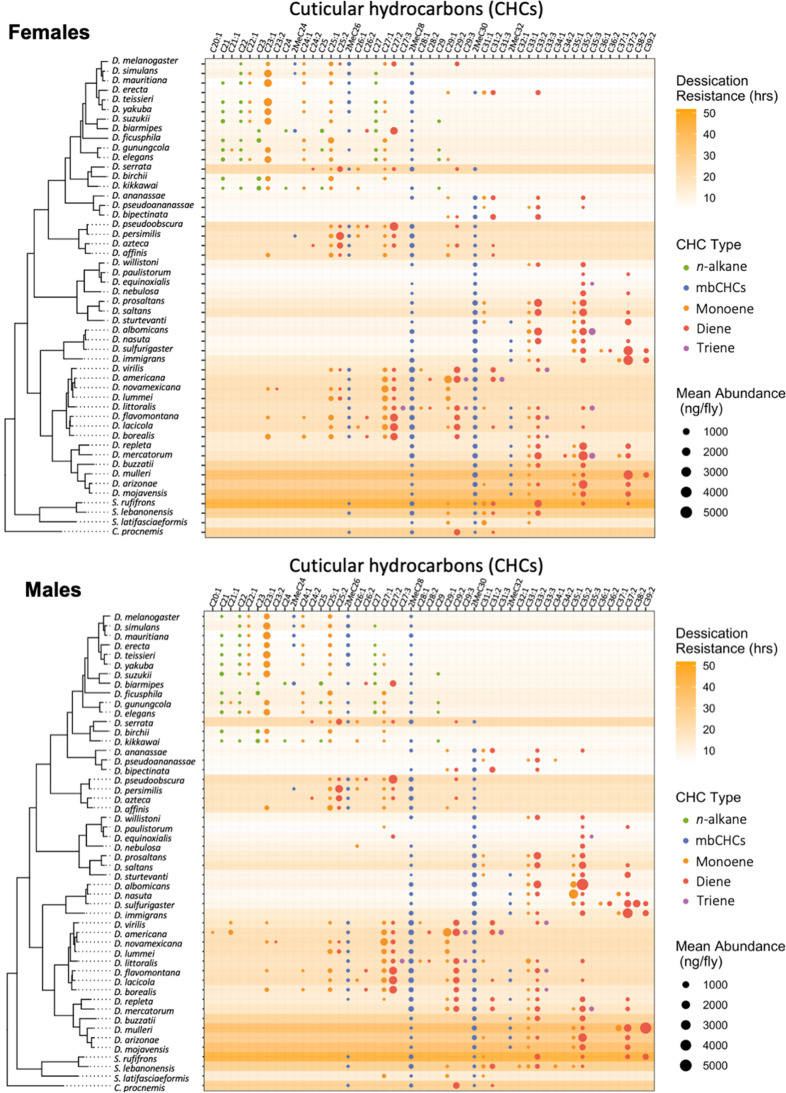
Similar articles
-
Rapid Changes in Composition and Contents of Cuticular Hydrocarbons in Sitobion avenae (Hemiptera: Aphididae) Clones Adapting to Desiccation Stress.J Econ Entomol. 2022 Apr 13;115(2):508-518. doi: 10.1093/jee/toab240. J Econ Entomol. 2022. PMID: 35022723
-
Quantitative genetic analysis suggests causal association between cuticular hydrocarbon composition and desiccation survival in Drosophila melanogaster.Heredity (Edinb). 2011 Jan;106(1):68-77. doi: 10.1038/hdy.2010.40. Epub 2010 Apr 14. Heredity (Edinb). 2011. PMID: 20389309 Free PMC article.
-
Desiccation Resistance and Micro-Climate Adaptation: Cuticular Hydrocarbon Signatures of Different Argentine Ant Supercolonies Across California.J Chem Ecol. 2018 Dec;44(12):1101-1114. doi: 10.1007/s10886-018-1029-y. Epub 2018 Nov 15. J Chem Ecol. 2018. PMID: 30430363
-
Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation.Heredity (Edinb). 2021 Feb;126(2):219-234. doi: 10.1038/s41437-020-00380-y. Epub 2020 Nov 2. Heredity (Edinb). 2021. PMID: 33139902 Free PMC article. Review.
-
Chemical Ecology, Biochemistry, and Molecular Biology of Insect Hydrocarbons.Annu Rev Entomol. 2021 Jan 7;66:45-60. doi: 10.1146/annurev-ento-031620-071754. Annu Rev Entomol. 2021. PMID: 33417824 Review.
Cited by
-
Comparisons of lifespan and stress resistance between sexes in Drosophila melanogaster.Heliyon. 2023 Jul 12;9(8):e18178. doi: 10.1016/j.heliyon.2023.e18178. eCollection 2023 Aug. Heliyon. 2023. PMID: 37576293 Free PMC article.
-
Latitudinal Clines in Climate and Sleep Patterns Shape Disease Outcomes in Drosophila melanogaster Infected by Metarhizium anisopliae.Ecol Evol. 2025 Feb 28;15(3):e71047. doi: 10.1002/ece3.71047. eCollection 2025 Mar. Ecol Evol. 2025. PMID: 40027417 Free PMC article.
-
Single-cell transcriptomics of clinical grade adipose-derived regenerative cells reveals consistency between donors independent of gender and BMI.Stem Cell Res Ther. 2025 Mar 5;16(1):109. doi: 10.1186/s13287-025-04234-4. Stem Cell Res Ther. 2025. PMID: 40038777 Free PMC article.
-
Regulatory changes in the fatty acid elongase eloF underlie the evolution of sex-specific pheromone profiles in Drosophila prolongata.BMC Biol. 2025 Apr 30;23(1):117. doi: 10.1186/s12915-025-02220-z. BMC Biol. 2025. PMID: 40307835 Free PMC article.
-
Cuticular hydrocarbons promote desiccation resistance by preventing transpiration in Drosophila melanogaster.J Exp Biol. 2024 Dec 1;227(23):jeb247752. doi: 10.1242/jeb.247752. Epub 2024 Nov 28. J Exp Biol. 2024. PMID: 39445981 Free PMC article.
References
-
- Belsley DA, Kuh E, Welsch RE. Regression diagnostics: Identifying influential data and sources of collinearity. John Wiley & Sons; 2005. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
