

Discovery and structure of a widespread bacterial ABC transporter specific for ergothioneine
- PMID: 36481738
- PMCID: PMC9732360
- DOI: 10.1038/s41467-022-35277-3
Discovery and structure of a widespread bacterial ABC transporter specific for ergothioneine
Abstract
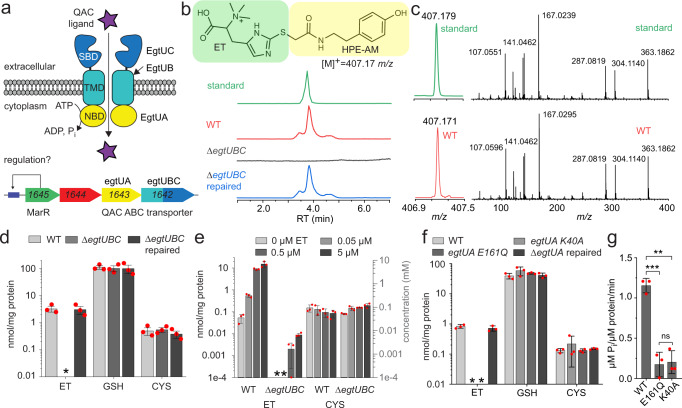
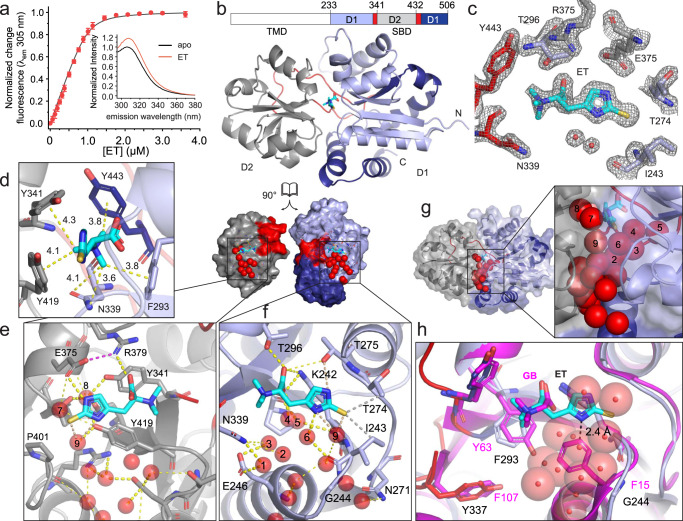
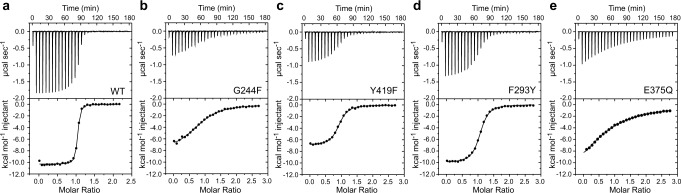
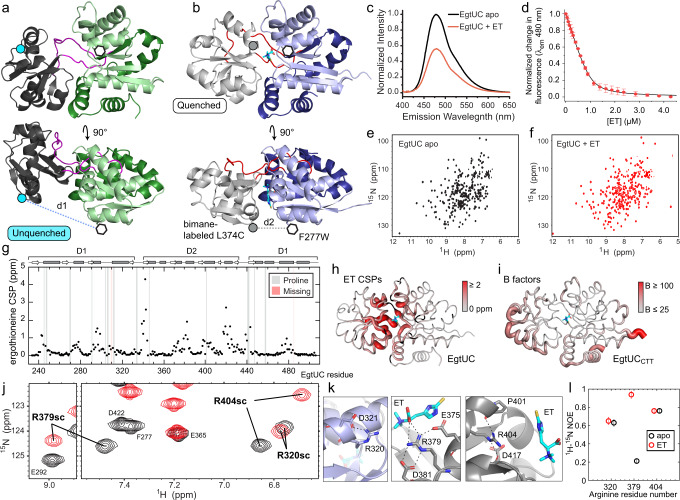
L-Ergothioneine (ET), the 2-thioimidazole derivative of trimethylhistidine, is biosynthesized by select fungi and bacteria, notably Mycobacterium tuberculosis, and functions as a scavenger of reactive oxygen species. The extent to which ET broadly functions in bacterial cells unable to synthesize it is unknown. Here we show that spd_1642-1643 in Streptococcus pneumoniae, a Gram-positive respiratory pathogen, encodes an ET uptake ATP-binding cassette (ABC) transporter, designated EgtU. The solute binding domain (SBD) of EgtU, EgtUC, binds ET with high affinity and exquisite specificity in a cleft between the two subdomains, with cation-π interactions engaging the betaine moiety and a network of water molecules that surround the thioimidazole ring. EgtU is highly conserved among known quaternary amine compound-specific transporters and widely distributed in Firmicutes, including the human pathogens Listeria monocytogenes, as BilEB, Enterococcus faecalis and Staphylococcus aureus. ET increases the chemical diversity of the low molecular weight thiol pool in Gram-positive human pathogens and may contribute to antioxidant defenses in the infected host.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
CH•••S hydrogen bonds drive molecular recognition of ergothioneine by the microbial transporter.bioRxiv [Preprint]. 2025 Jul 31:2025.07.28.667264. doi: 10.1101/2025.07.28.667264. bioRxiv. 2025. PMID: 40766558 Free PMC article. Preprint.
-
Expression and purification of the intact bacterial ergothioneine transporter EgtU.Protein Expr Purif. 2025 Mar;227:106633. doi: 10.1016/j.pep.2024.106633. Epub 2024 Nov 20. Protein Expr Purif. 2025. PMID: 39577818
-
Functional diversity of tandem substrate-binding domains in ABC transporters from pathogenic bacteria.Structure. 2013 Oct 8;21(10):1879-88. doi: 10.1016/j.str.2013.07.020. Epub 2013 Aug 29. Structure. 2013. PMID: 23994008
-
The ergothioneine transporter controls and indicates ergothioneine activity--a review.Prev Med. 2012 May;54 Suppl:S71-4. doi: 10.1016/j.ypmed.2011.12.001. Epub 2011 Dec 13. Prev Med. 2012. PMID: 22182480 Review.
-
Ergothioneine, an adaptive antioxidant for the protection of injured tissues? A hypothesis.Biochem Biophys Res Commun. 2016 Feb 5;470(2):245-250. doi: 10.1016/j.bbrc.2015.12.124. Epub 2016 Jan 6. Biochem Biophys Res Commun. 2016. PMID: 26772879 Review.
Cited by
-
Understanding mechanisms of antioxidant action in health and disease.Nat Rev Mol Cell Biol. 2024 Jan;25(1):13-33. doi: 10.1038/s41580-023-00645-4. Epub 2023 Sep 15. Nat Rev Mol Cell Biol. 2024. PMID: 37714962 Review.
-
CH•••S hydrogen bonds drive molecular recognition of ergothioneine by the microbial transporter.bioRxiv [Preprint]. 2025 Jul 31:2025.07.28.667264. doi: 10.1101/2025.07.28.667264. bioRxiv. 2025. PMID: 40766558 Free PMC article. Preprint.
-
An affinity-based depletion strategy for evaluating the effects of ergothioneine on bacterial physiology.Cell Chem Biol. 2025 Mar 20;32(3):486-497.e7. doi: 10.1016/j.chembiol.2025.02.004. Epub 2025 Mar 10. Cell Chem Biol. 2025. PMID: 40068683
-
Impact of Difluoromethylornithine and AMXT 1501 on Gene Expression and Capsule Regulation in Streptococcus pneumoniae.Biomolecules. 2024 Feb 2;14(2):178. doi: 10.3390/biom14020178. Biomolecules. 2024. PMID: 38397415 Free PMC article.
-
Bacterial cell volume regulation and the importance of cyclic di-AMP.Microbiol Mol Biol Rev. 2024 Jun 27;88(2):e0018123. doi: 10.1128/mmbr.00181-23. Epub 2024 Jun 10. Microbiol Mol Biol Rev. 2024. PMID: 38856222 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
