

Autoinhibition and activation mechanisms revealed by the triangular-shaped structure of myosin Va
- PMID: 36490350
- PMCID: PMC9733927
- DOI: 10.1126/sciadv.add4187
Autoinhibition and activation mechanisms revealed by the triangular-shaped structure of myosin Va
Abstract
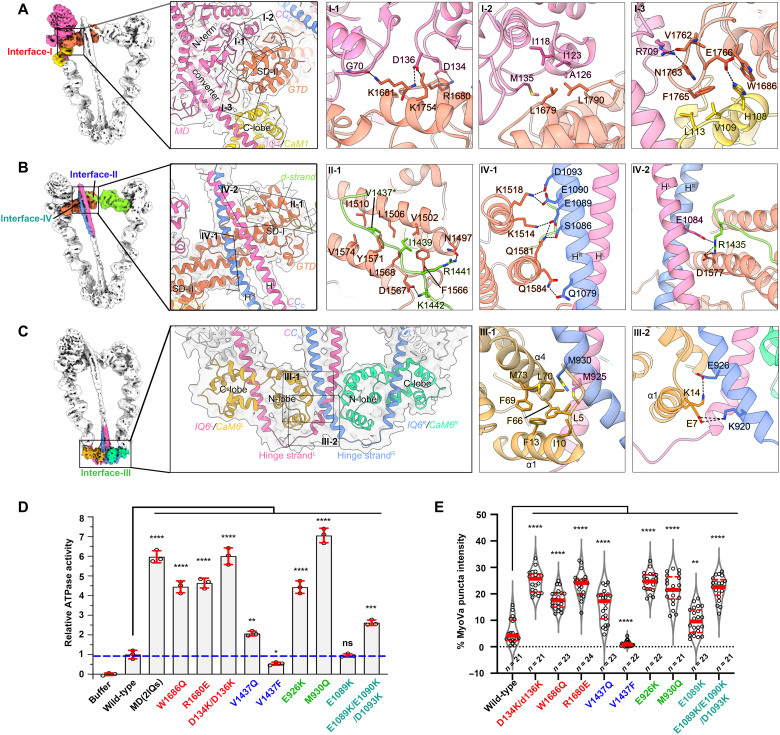
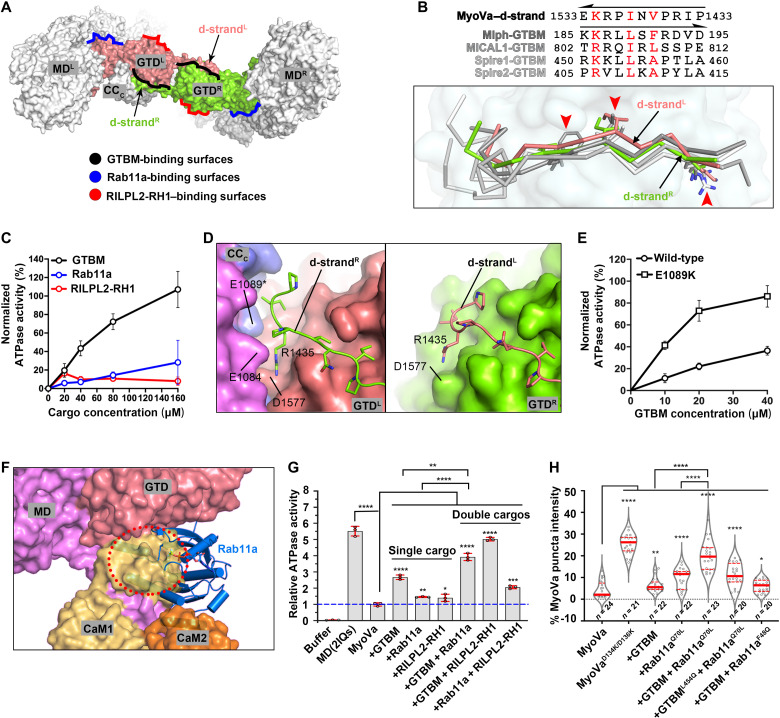
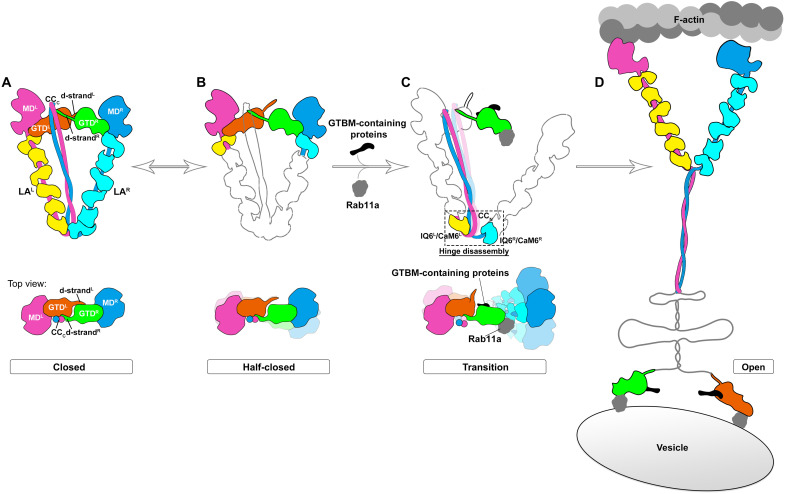
As the prototype of unconventional myosin motor family, myosin Va (MyoVa) transport cellular cargos along actin filaments in diverse cellular processes. The off-duty MyoVa adopts a closed and autoinhibited state, which can be relieved by cargo binding. The molecular mechanisms governing the autoinhibition and activation of MyoVa remain unclear. Here, we report the cryo-electron microscopy structure of the two full-length, closed MyoVa heavy chains in complex with 12 calmodulin light chains at 4.78-Å resolution. The MyoVa adopts a triangular structure with multiple intra- and interpolypeptide chain interactions in establishing the closed state with cargo binding and adenosine triphosphatase activity inhibited. Structural, biochemical, and cellular analyses uncover an asymmetric autoinhibition mechanism, in which the cargo-binding sites in the two MyoVa heavy chains are differently protected. Thus, specific and efficient MyoVa activation requires coincident binding of multiple cargo adaptors, revealing an intricate and elegant activity regulation of the motor in response to cargos.
Figures



References
-
- N. Hirokawa, R. Takemura,Molecular motors and mechanisms of directional transport in neurons. Nat. Rev. Neurosci. 6,201–214 (2005). - PubMed
-
- N. Hirokawa, S. Niwa, Y. Tanaka,Molecular motors in neurons: Transport mechanisms and roles in brain function, development, and disease. Neuron 68,610–638 (2010). - PubMed
-
- M. Kneussel, W. Wagner,Myosin motors at neuronal synapses: Drivers of membrane transport and actin dynamics. Nat. Rev. Neurosci. 14,233–247 (2013). - PubMed
-
- R. D. Vale,The molecular motor toolbox for intracellular transport. Cell 112,467–480 (2003). - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
