

The histone methyltransferase MLL1/KMT2A in monocytes drives coronavirus-associated coagulopathy and inflammation
- PMID: 36493338
- PMCID: PMC9743412
- DOI: 10.1182/blood.2022015917
The histone methyltransferase MLL1/KMT2A in monocytes drives coronavirus-associated coagulopathy and inflammation
Abstract
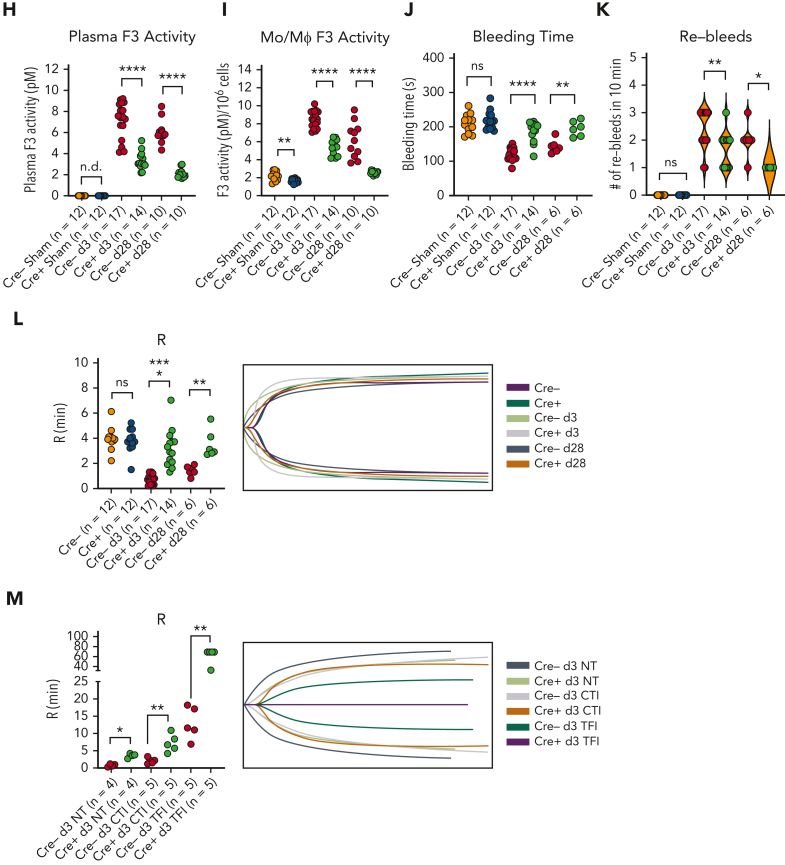
Coronavirus-associated coagulopathy (CAC) is a morbid and lethal sequela of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection. CAC results from a perturbed balance between coagulation and fibrinolysis and occurs in conjunction with exaggerated activation of monocytes/macrophages (MO/Mφs), and the mechanisms that collectively govern this phenotype seen in CAC remain unclear. Here, using experimental models that use the murine betacoronavirus MHVA59, a well-established model of SARS-CoV-2 infection, we identify that the histone methyltransferase mixed lineage leukemia 1 (MLL1/KMT2A) is an important regulator of MO/Mφ expression of procoagulant and profibrinolytic factors such as tissue factor (F3; TF), urokinase (PLAU), and urokinase receptor (PLAUR) (herein, "coagulopathy-related factors") in noninfected and infected cells. We show that MLL1 concurrently promotes the expression of the proinflammatory cytokines while suppressing the expression of interferon alfa (IFN-α), a well-known inducer of TF and PLAUR. Using in vitro models, we identify MLL1-dependent NF-κB/RelA-mediated transcription of these coagulation-related factors and identify a context-dependent, MLL1-independent role for RelA in the expression of these factors in vivo. As functional correlates for these findings, we demonstrate that the inflammatory, procoagulant, and profibrinolytic phenotypes seen in vivo after coronavirus infection were MLL1-dependent despite blunted Ifna induction in MO/Mφs. Finally, in an analysis of SARS-CoV-2 positive human samples, we identify differential upregulation of MLL1 and coagulopathy-related factor expression and activity in CD14+ MO/Mφs relative to noninfected and healthy controls. We also observed elevated plasma PLAU and TF activity in COVID-positive samples. Collectively, these findings highlight an important role for MO/Mφ MLL1 in promoting CAC and inflammation.
© 2023 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: M.H. is a consultant and equity holder for Veralox Therapeutics and a consultant for Cereno Scientific, which has an option to license platelet inhibitory compounds from the University of Michigan. The remaining authors declare no competing financial interests.
Figures











Comment in
-
MLL1 is central to macrophage-mediated inflammation.Blood. 2023 Feb 16;141(7):687-689. doi: 10.1182/blood.2022019181. Blood. 2023. PMID: 36795448 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK131782/DK/NIDDK NIH HHS/United States
- R01 DK127531/DK/NIDDK NIH HHS/United States
- R21 TR003185/TR/NCATS NIH HHS/United States
- R35 HL167143/HL/NHLBI NIH HHS/United States
- R35 HL144481/HL/NHLBI NIH HHS/United States
- ZIA HL006262/ImNIH/Intramural NIH HHS/United States
- R01 HL144550/HL/NHLBI NIH HHS/United States
- ZIA HL006263/ImNIH/Intramural NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- R01 DK124290/DK/NIDDK NIH HHS/United States
- ZIA HL006267/ImNIH/Intramural NIH HHS/United States
- R35 GM131835/GM/NIGMS NIH HHS/United States
- K08 HL155408/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
