

Fibroblast inflammatory priming determines regenerative versus fibrotic skin repair in reindeer
- PMID: 36493752
- PMCID: PMC9888357
- DOI: 10.1016/j.cell.2022.11.004
Fibroblast inflammatory priming determines regenerative versus fibrotic skin repair in reindeer
Abstract
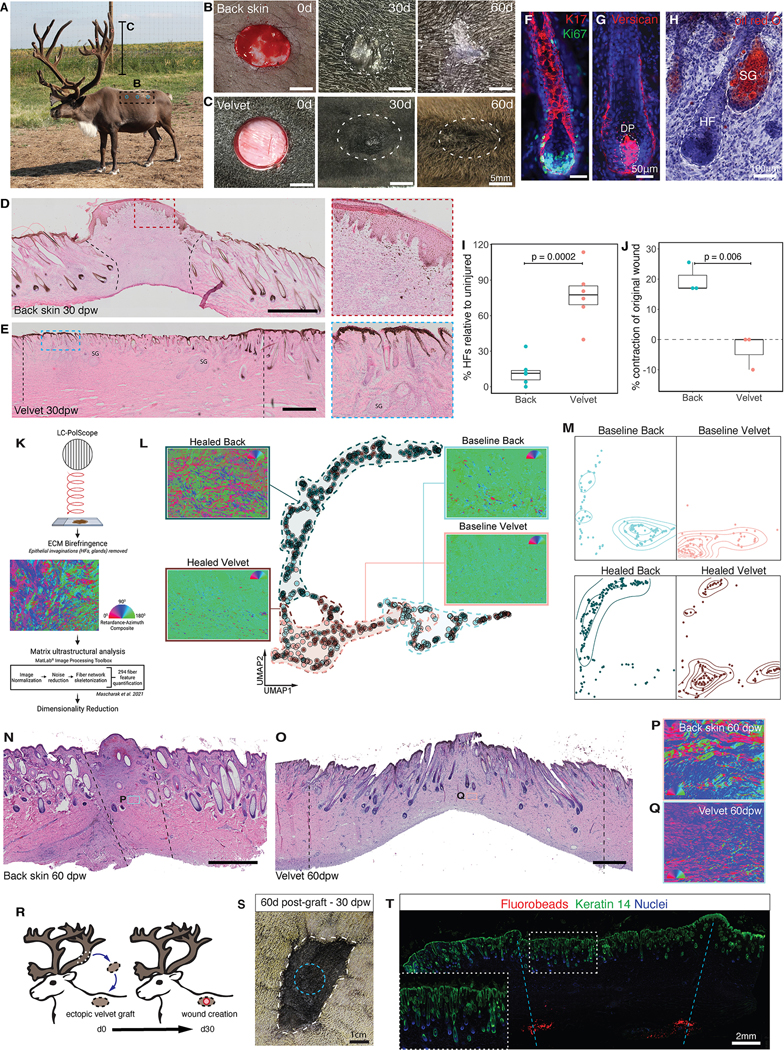
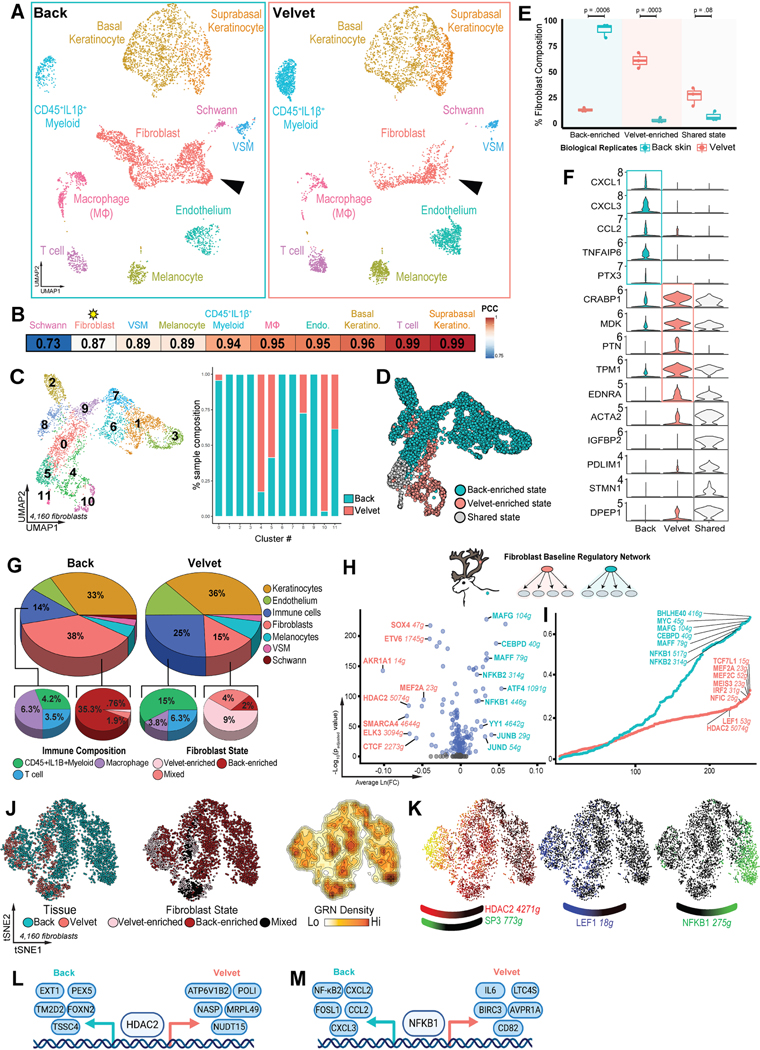
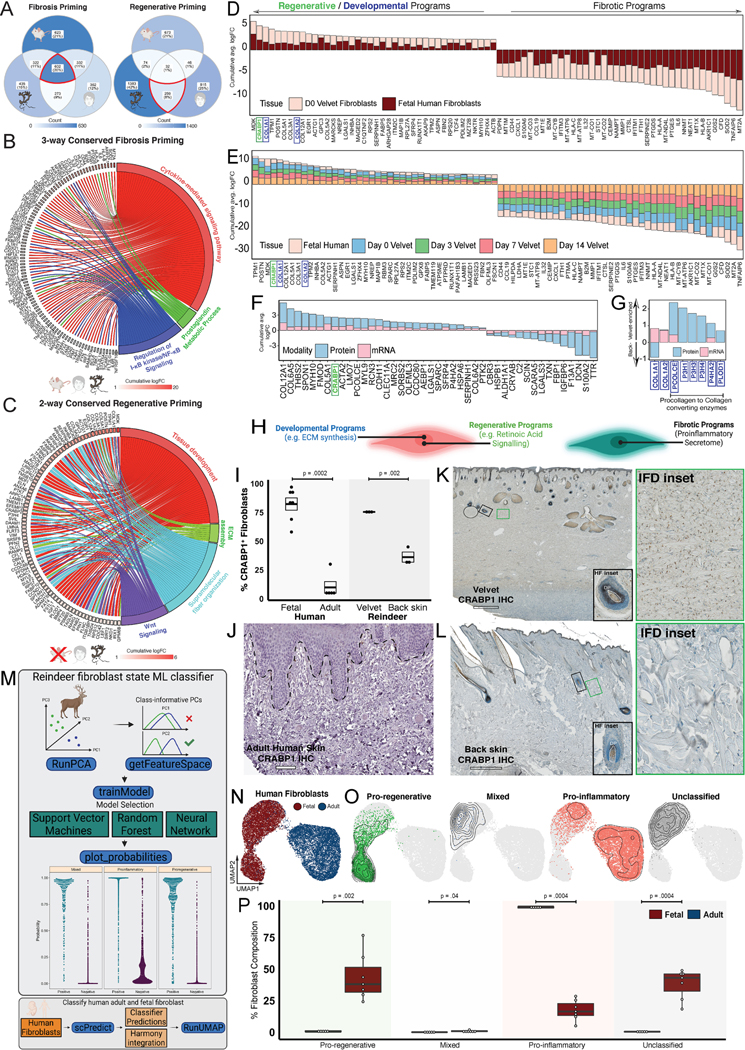
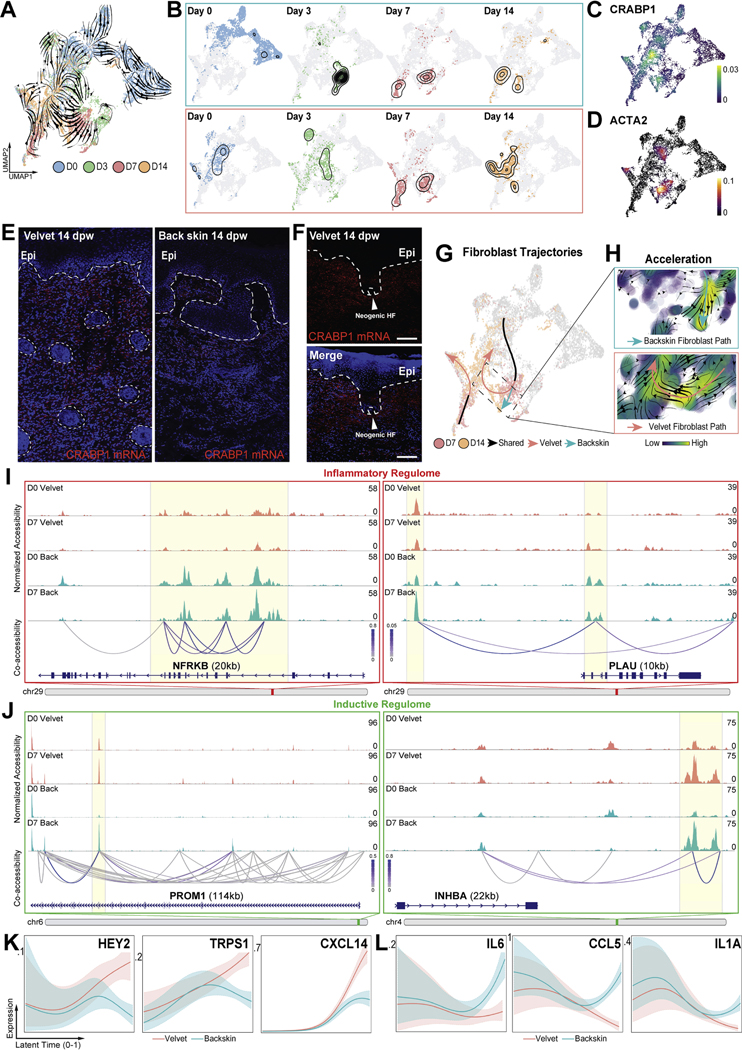
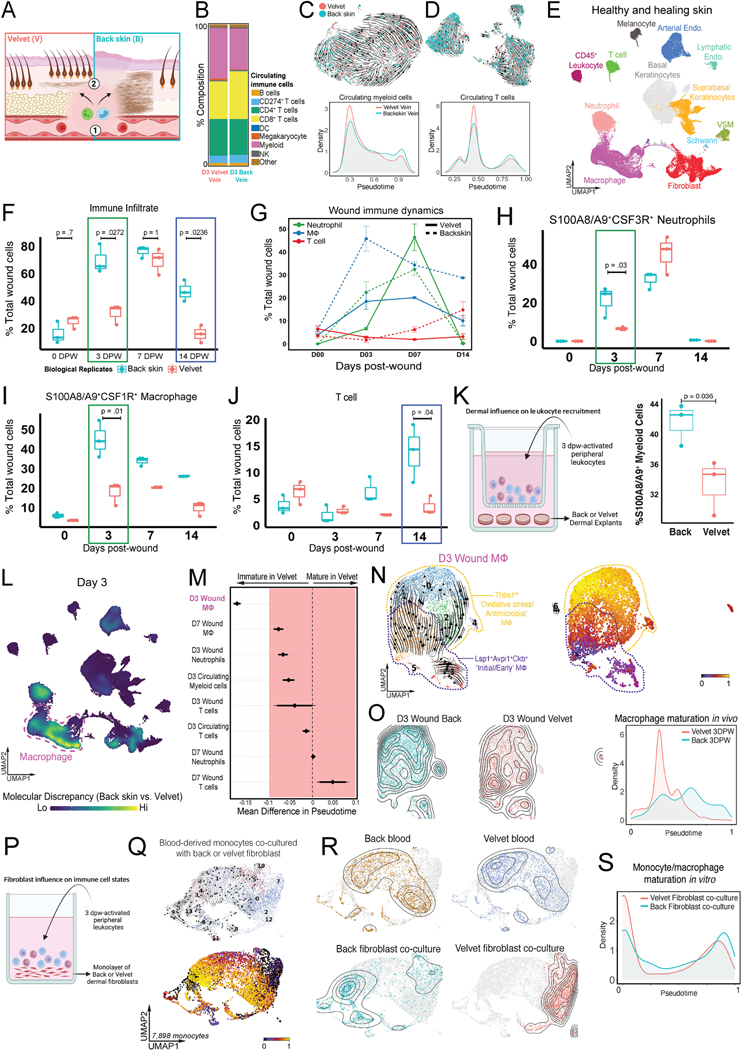
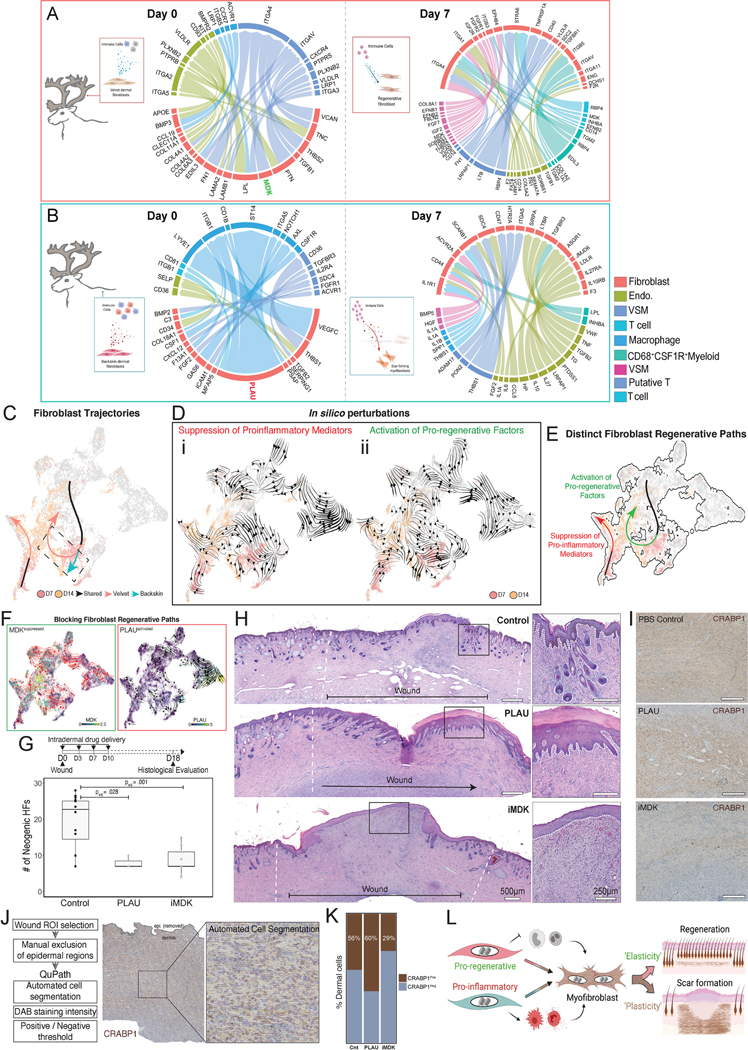
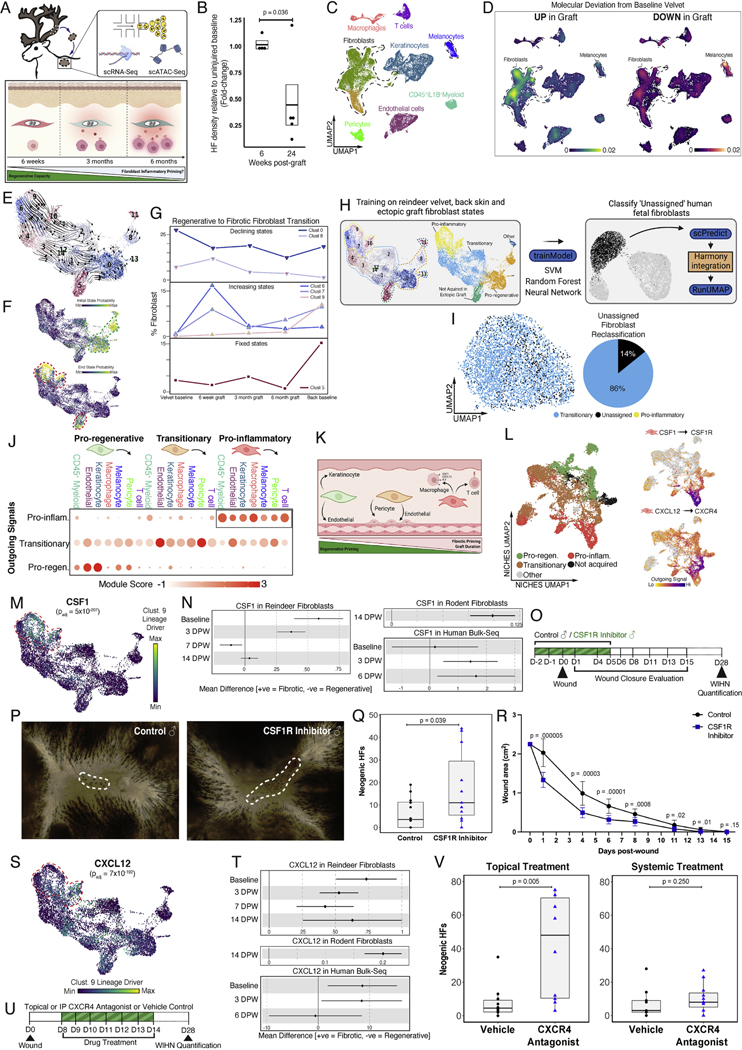
Adult mammalian skin wounds heal by forming fibrotic scars. We report that full-thickness injuries of reindeer antler skin (velvet) regenerate, whereas back skin forms fibrotic scar. Single-cell multi-omics reveal that uninjured velvet fibroblasts resemble human fetal fibroblasts, whereas back skin fibroblasts express inflammatory mediators mimicking pro-fibrotic adult human and rodent fibroblasts. Consequently, injury elicits site-specific immune responses: back skin fibroblasts amplify myeloid infiltration and maturation during repair, whereas velvet fibroblasts adopt an immunosuppressive phenotype that restricts leukocyte recruitment and hastens immune resolution. Ectopic transplantation of velvet to scar-forming back skin is initially regenerative, but progressively transitions to a fibrotic phenotype akin to the scarless fetal-to-scar-forming transition reported in humans. Skin regeneration is diminished by intensifying, or enhanced by neutralizing, these pathologic fibroblast-immune interactions. Reindeer represent a powerful comparative model for interrogating divergent wound healing outcomes, and our results nominate decoupling of fibroblast-immune interactions as a promising approach to mitigate scar.
Keywords: fetal human fibroblast; fibroblast; immune modulation; inflammation; inflammatory priming; myeloid cell maturation; reindeer; scar; skin regeneration; stromal-immune crosstalk; wound healing.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Reindeer antlers model skin regeneration.Nat Rev Immunol. 2023 Feb;23(2):71. doi: 10.1038/s41577-022-00828-8. Nat Rev Immunol. 2023. PMID: 36513845 No abstract available.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
