

RNA degradation eliminates developmental transcripts during murine embryonic stem cell differentiation via CAPRIN1-XRN2
- PMID: 36495875
- PMCID: PMC9796812
- DOI: 10.1016/j.devcel.2022.11.014
RNA degradation eliminates developmental transcripts during murine embryonic stem cell differentiation via CAPRIN1-XRN2
Abstract
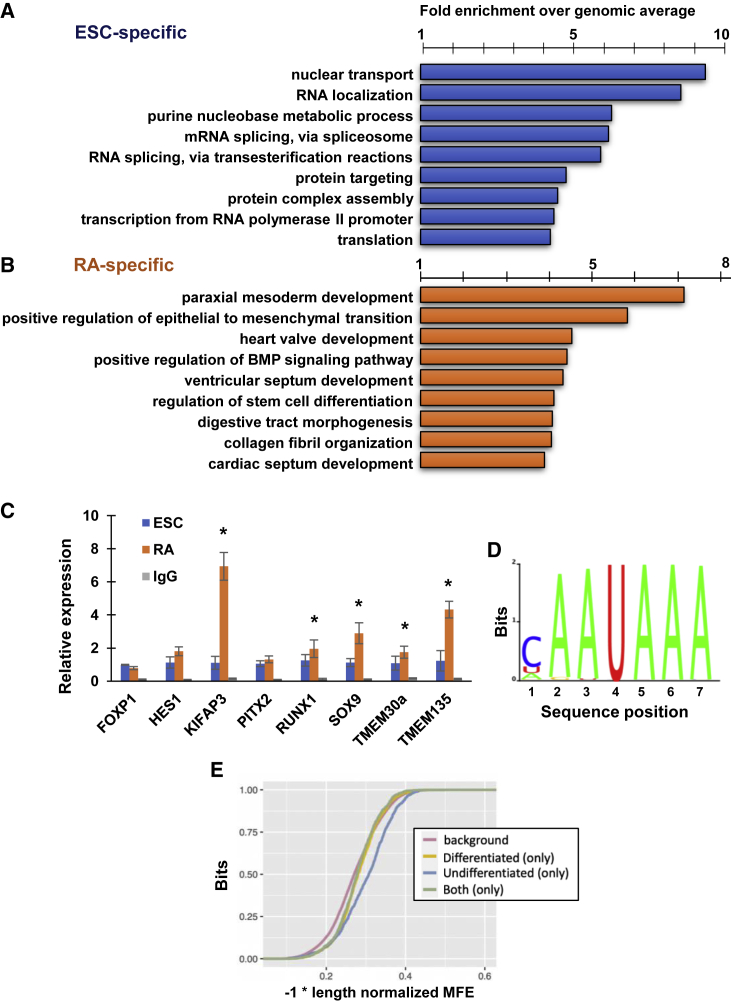
Embryonic stem cells (ESCs) are self-renewing and pluripotent. In recent years, factors that control pluripotency, mostly nuclear, have been identified. To identify non-nuclear regulators of ESCs, we screened an endogenously labeled fluorescent fusion-protein library in mouse ESCs. One of the more compelling hits was the cell-cycle-associated protein 1 (CAPRIN1). CAPRIN1 knockout had little effect in ESCs, but it significantly altered differentiation and gene expression programs. Using RIP-seq and SLAM-seq, we found that CAPRIN1 associates with, and promotes the degradation of, thousands of RNA transcripts. CAPRIN1 interactome identified XRN2 as the likely ribonuclease. Upon early ESC differentiation, XRN2 is located in the nucleus and colocalizes with CAPRIN1 in small RNA granules in a CAPRIN1-dependent manner. We propose that CAPRIN1 regulates an RNA degradation pathway operating during early ESC differentiation, thus eliminating undesired spuriously transcribed transcripts in ESCs.
Keywords: CAPRIN1; RNA degradation; RNA stability; XRN2; differentiation; embryonic stem cells; pluripotency; stress granules.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Similar articles
-
BTG/Tob family members Tob1 and Tob2 inhibit proliferation of mouse embryonic stem cells via Id3 mRNA degradation.Biochem Biophys Res Commun. 2015 Jul 3;462(3):208-14. doi: 10.1016/j.bbrc.2015.04.117. Epub 2015 May 5. Biochem Biophys Res Commun. 2015. PMID: 25951976
-
Perturbed differentiation of murine embryonic stem cells upon Pelota deletion due to dysregulated FOXO1/β-catenin signaling.FEBS J. 2021 May;288(10):3317-3329. doi: 10.1111/febs.15643. Epub 2020 Dec 17. FEBS J. 2021. PMID: 33245852
-
Allele-specific RNA-seq expression profiling of imprinted genes in mouse isogenic pluripotent states.Epigenetics Chromatin. 2019 Feb 15;12(1):14. doi: 10.1186/s13072-019-0259-8. Epigenetics Chromatin. 2019. PMID: 30767785 Free PMC article.
-
Rbm24 Regulates Alternative Splicing Switch in Embryonic Stem Cell Cardiac Lineage Differentiation.Stem Cells. 2016 Jul;34(7):1776-89. doi: 10.1002/stem.2366. Epub 2016 Mar 28. Stem Cells. 2016. PMID: 26990106
-
The association and clinical relevance of phase-separating protein CAPRIN1 with noncoding RNA.Cell Stress Chaperones. 2023 Mar;28(2):125-132. doi: 10.1007/s12192-023-01320-5. Epub 2023 Jan 11. Cell Stress Chaperones. 2023. PMID: 36627508 Free PMC article. Review.
Cited by
-
An antibody-dependent cellular phagocytosis-related gene signature predicts survival and response to immunotherapy in stomach adenocarcinoma.Medicine (Baltimore). 2025 Apr 4;104(14):e42079. doi: 10.1097/MD.0000000000042079. Medicine (Baltimore). 2025. PMID: 40193680 Free PMC article.
-
Long noncoding RNA GDIL acts as a scaffold for CHAC1 and XRN2 to promote platinum resistance of colorectal cancer through inhibition of glutathione degradation.Cell Death Dis. 2025 Feb 1;16(1):62. doi: 10.1038/s41419-025-07374-w. Cell Death Dis. 2025. PMID: 39893168 Free PMC article.
-
Characterization of the long noncoding RNA transcriptome in human preimplantation embryo development.J Assist Reprod Genet. 2023 Dec;40(12):2913-2923. doi: 10.1007/s10815-023-02951-4. Epub 2023 Sep 29. J Assist Reprod Genet. 2023. PMID: 37770818 Free PMC article.
-
β-propeller protein-associated neurodegeneration protein WDR45 regulates stress granule disassembly via phase separation with Caprin-1.Nat Commun. 2025 Jun 5;16(1):5227. doi: 10.1038/s41467-025-60583-x. Nat Commun. 2025. PMID: 40473629 Free PMC article.
-
Clearing the slate: RNA turnover to enable cell state switching?Development. 2023 Oct 1;150(19):dev202084. doi: 10.1242/dev.202084. Epub 2023 Oct 12. Development. 2023. PMID: 37831057 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
