

CRL2ZER1/ZYG11B recognizes small N-terminal residues for degradation
- PMID: 36496439
- PMCID: PMC9741652
- DOI: 10.1038/s41467-022-35169-6
CRL2ZER1/ZYG11B recognizes small N-terminal residues for degradation
Abstract
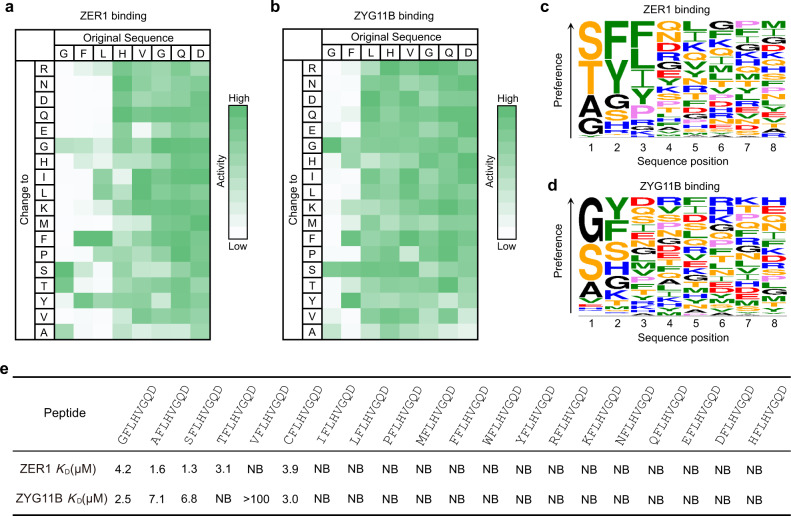
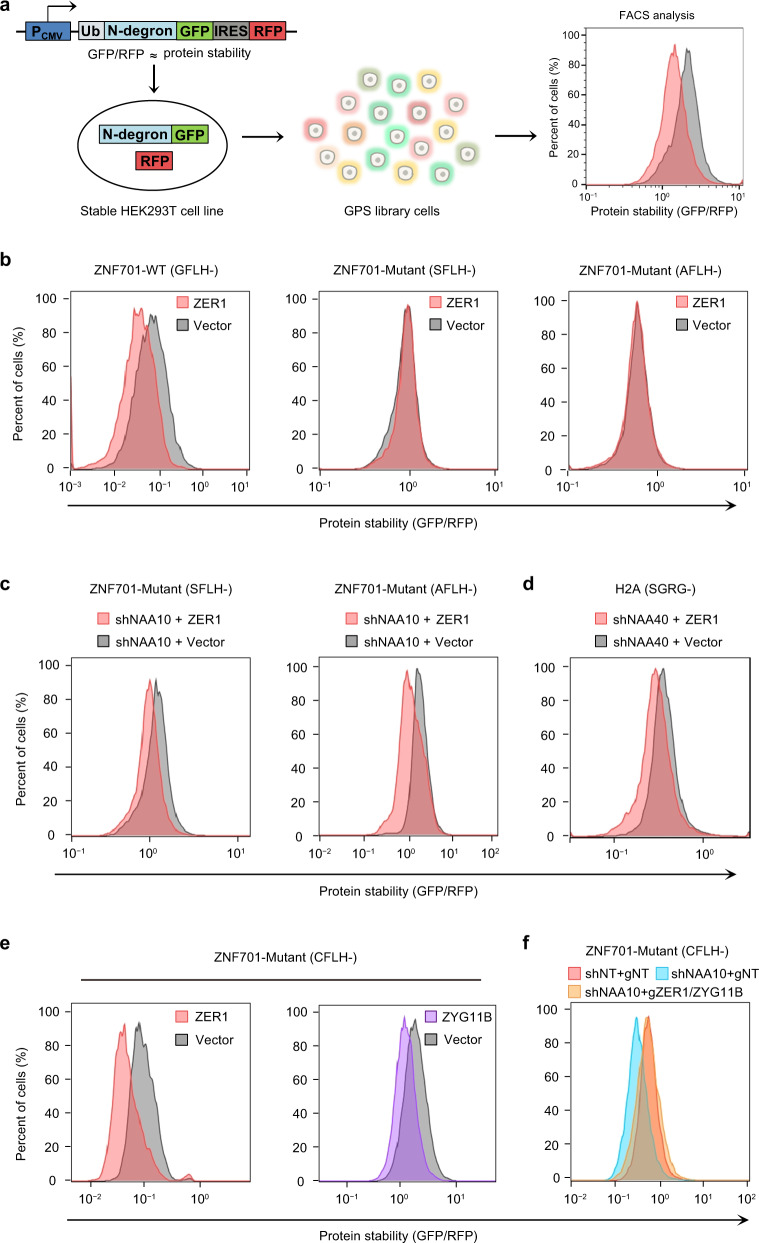
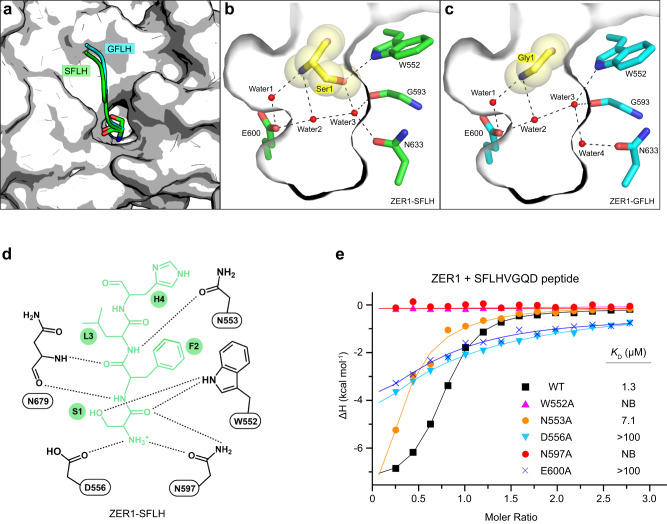
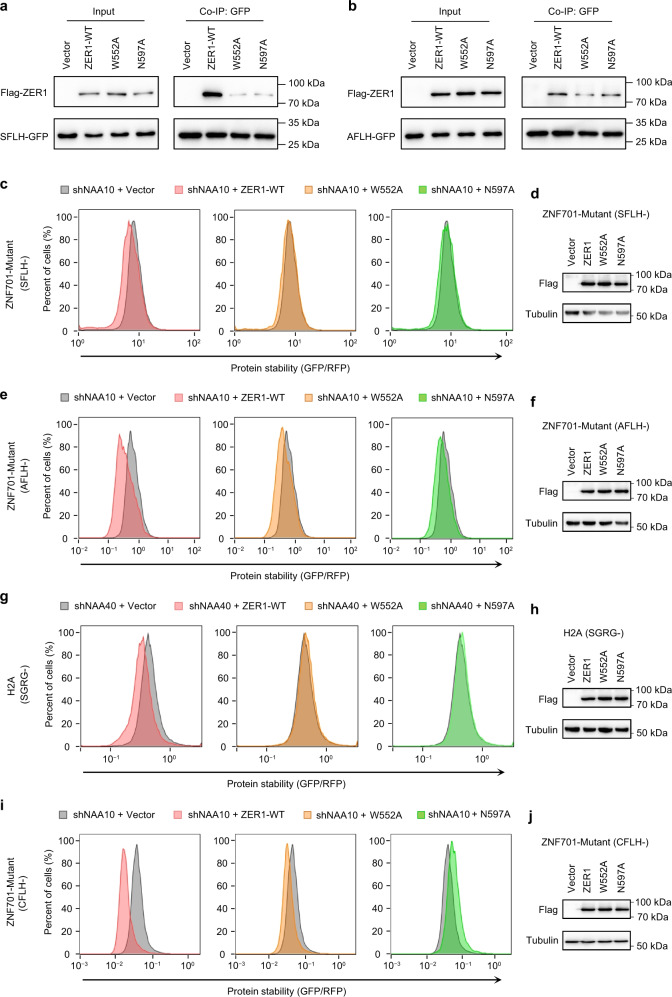
N-degron pathway plays an important role in the protein quality control and maintenance of cellular protein homeostasis. ZER1 and ZYG11B, the substrate receptors of the Cullin 2-RING E3 ubiquitin ligase (CRL2), recognize N-terminal (Nt) glycine degrons and participate in the Nt-myristoylation quality control through the Gly/N-degron pathway. Here we show that ZER1 and ZYG11B can also recognize small Nt-residues other than glycine. Specifically, ZER1 binds better to Nt-Ser, -Ala, -Thr and -Cys than to -Gly, while ZYG11B prefers Nt-Gly but also has the capacity to recognize Nt-Ser, -Ala and -Cys in vitro. We found that Nt-Ser, -Ala and -Cys undergo Nt-acetylation catalyzed by Nt-acetyltransferase (NAT), thereby shielding them from recognition by ZER1/ZYG11B in cells. Instead, ZER1/ZYG11B readily targets a selection of small Nt-residues lacking Nt-acetylation for degradation in NAT-deficient cells, implicating its role in the Nt-acetylation quality control. Furthermore, we present the crystal structures of ZER1 and ZYG11B bound to various small Nt-residues and uncover the molecular mechanism of non-acetylated substrate recognition by ZER1 and ZYG11B.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
