

Cryo-electron tomography reveals structural insights into the membrane remodeling mode of dynamin-like EHD filaments
- PMID: 36496453
- PMCID: PMC9741607
- DOI: 10.1038/s41467-022-35164-x
Cryo-electron tomography reveals structural insights into the membrane remodeling mode of dynamin-like EHD filaments
Abstract
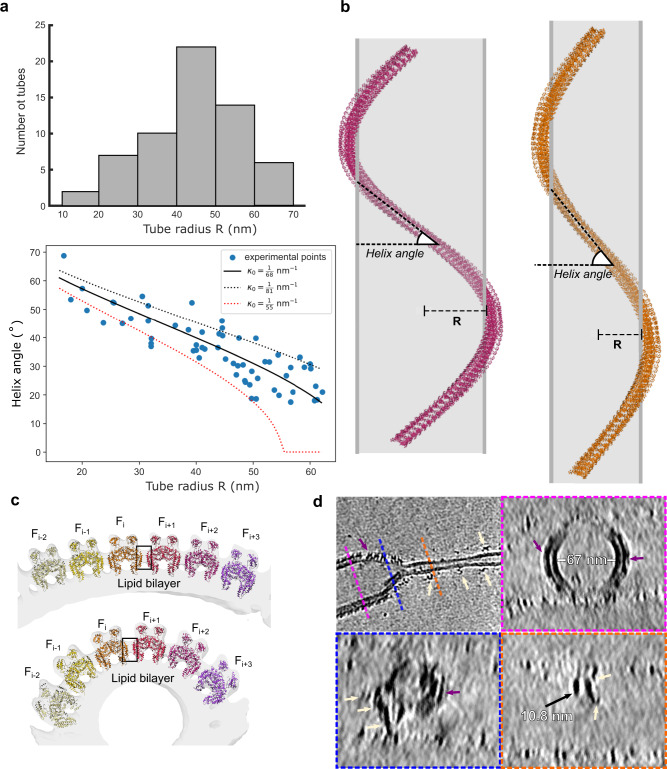
Eps15-homology domain containing proteins (EHDs) are eukaryotic, dynamin-related ATPases involved in cellular membrane trafficking. They oligomerize on membranes into filaments that induce membrane tubulation. While EHD crystal structures in open and closed conformations were previously reported, little structural information is available for the membrane-bound oligomeric form. Consequently, mechanistic insights into the membrane remodeling mechanism have remained sparse. Here, by using cryo-electron tomography and subtomogram averaging, we determined structures of nucleotide-bound EHD4 filaments on membrane tubes of various diameters at an average resolution of 7.6 Å. Assembly of EHD4 is mediated via interfaces in the G-domain and the helical domain. The oligomerized EHD4 structure resembles the closed conformation, where the tips of the helical domains protrude into the membrane. The variation in filament geometry and tube radius suggests a spontaneous filament curvature of approximately 1/70 nm-1. Combining the available structural and functional data, we suggest a model for EHD-mediated membrane remodeling.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
