

Delivery of Active AKT1 to Human Cells
- PMID: 36497091
- PMCID: PMC9738475
- DOI: 10.3390/cells11233834
Delivery of Active AKT1 to Human Cells
Abstract
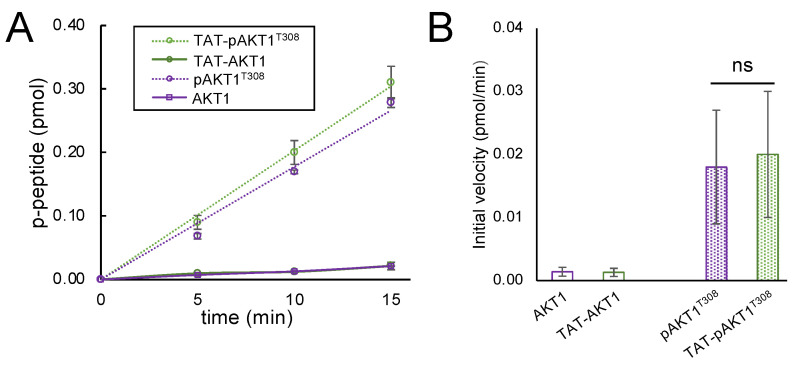
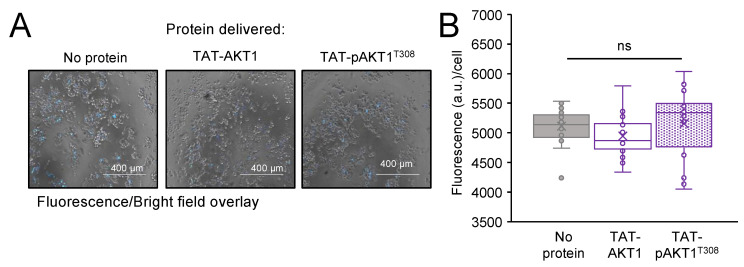
Protein kinase B (AKT1) is a serine/threonine kinase and central transducer of cell survival pathways. Typical approaches to study AKT1 biology in cells rely on growth factor or insulin stimulation that activates AKT1 via phosphorylation at two key regulatory sites (Thr308, Ser473), yet cell stimulation also activates many other kinases. To produce cells with specific AKT1 activity, we developed a novel system to deliver active AKT1 to human cells. We recently established a method to produce AKT1 phospho-variants from Escherichia coli with programmed phosphorylation. Here, we fused AKT1 with an N-terminal cell penetrating peptide tag derived from the human immunodeficiency virus trans-activator of transcription (TAT) protein. The TAT-tag did not alter AKT1 kinase activity and was necessary and sufficient to rapidly deliver AKT1 protein variants that persisted in human cells for 24 h without the need to use transfection reagents. TAT-pAKT1T308 induced selective phosphorylation of the known AKT1 substrate GSK-3α, but not GSK-3β, and downstream stimulation of the AKT1 pathway as evidenced by phosphorylation of ribosomal protein S6 at Ser240/244. The data demonstrate efficient delivery of AKT1 with programmed phosphorylation to human cells, thus establishing a cell-based model system to investigate signaling that is dependent on AKT1 activity.
Keywords: cell penetrating peptide; cellular signaling; kinase; phosphoinositide-dependent kinase (PDK1); protein kinase B (AKT1); recombinant protein; trans-activator of transcription (TAT).
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Karege F., Perroud N., Schürhoff F., Meary A., Marillier G., Burkhardt S., Ballmann E., Fernandez R., Jamain S., Leboyer M. Association of AKT1 gene variants and protein expression in both schizophrenia and bipolar disorder. Genes Brain Behav. 2010;9:503–511. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
